
Roles of RNA Sensors in Host Innate Response to Influenza Virus and Coronavirus Infections


1
Key Laboratory of Animal Epidemiology of the Ministry of Agriculture, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
2
Department of Preventive Veterinary Medicine, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(15), 8285; https://doi.org/10.3390/ijms23158285
Submission received: 31 May 2022
/
Revised: 22 July 2022
/
Accepted: 23 July 2022
/
Published: 27 July 2022
(This article belongs to the Special Issue Host-Microbe Interaction 2022)
Abstract
:Influenza virus and coronavirus are two important respiratory viruses, which often cause serious respiratory diseases in humans and animals after infection. In recent years, highly pathogenic avian influenza virus (HPAIV) and SARS-CoV-2 have become major pathogens causing respiratory diseases in humans. Thus, an in-depth understanding of the relationship between viral infection and host innate immunity is particularly important to the stipulation of effective control strategies. As the first line of defense against pathogens infection, innate immunity not only acts as a natural physiological barrier, but also eliminates pathogens through the production of interferon (IFN), the formation of inflammasomes, and the production of pro-inflammatory cytokines. In this process, the recognition of viral pathogen-associated molecular patterns (PAMPs) by host pattern recognition receptors (PRRs) is the initiation and the most important part of the innate immune response. In this review, we summarize the roles of RNA sensors in the host innate immune response to influenza virus and coronavirus infections in different species, with a particular focus on innate immune recognition of viral nucleic acids in host cells, which will help to develop an effective strategy for the control of respiratory infectious diseases.
1. Introduction
Innate immunity is the natural resistance of the host against pathogens, including viruses, bacteria, fungi, and others. The specific components of pathogens including nucleic acids, proteins, peptidoglycan, and so on, which are called pathogen-associated molecular patterns (PAMPs), can be recognized by the sensors of host cells that are called pattern recognition receptors (PRRs) to initiate the innate immune response. Up to now, at least five groups of PRRs have been identified, including toll-like receptors (TLRs), nucleotide-binding oligomerization domain-like receptors (NLRs), retinoic acid-inducible gene I-like helicases receptors (RLRs), c-type lectin receptors (CLRs), and DNA sensors including AIM2-like receptors (ALRs) and Cyclic-GMP-AMP Synthase (cGAS). These PRRs have different structures and are located differently in host cells. Upon engagement of PAMPs by host PRRs, the signal-transducing pathway is initiated with the recruitment of adaptors to the activating domains of PRRs, ultimately eliciting the innate immune response.
As the transmembrane proteins, TLRs recognize the nucleic acids, lipoproteins, and lipopolysaccharides (LPS) of pathogens [1,2,3,4]. The engagement of TLRs with PAMPs leads to the recruitment and activation of downstream adapters, such as myeloid differentiation factor 88 (MyD88), TIR domain-containing adapter-inducing interferon-β (TRIF), and TIR domain-containing adapter protein (TIRAP). These adapters are involved in NF-κB, IRF, or AP-1 signaling pathways to trigger innate immune responses [5,6,7]. RLRs, mainly existing in the cytoplasm, consisting of several members, such as retinoic acid-inducible gene I (RIG-I), melanoma differentiation-associated gene 5 (MDA5), and laboratory of genetics and physiology 2 (LGP2), sensing the pathogenic double-stranded RNA (dsRNA)to activate the mitochondrial antiviral signaling protein (MVAS) through the unique CARD domain, and initiating NF-κB and IRFs signaling pathways [8,9,10,11,12,13]. NLRs, a member of PRRs in the cytoplasm, primarily sense certain components of bacteria. Among them, the most important representatives are NOD1 and NOD2. Once the bacterial peptidoglycan (PGN) is recognized by NOD1/NOD2, the NF-κB and IRFs signaling pathways are subsequently activated by recruiting and activating the receptor interacting protein 2 (RIP2), triggering an innate immune response [14,15,16]. The engagement of NLRs by PAMPs of bacteria or damage-associated molecular patterns (DAMPs) also induces the formation of inflammasomes, which activate pro-caspase-1 to cleave the gasdermin family proteins, resulting in pyroptosis and inflammatory response [17,18,19]. CLRs are cell membrane receptors that recognize fungi [20]. Once sensing appropriate ligands, CLRs will act on MALT1/Bcl-10 by activating spleen tyrosine kinase (Syk) through its ITAM domain, and finally activate NF-κB and AP-1 signaling pathways [21]. In addition, the activated Syk can slao induce the innate immune response by activating the NF-AT signaling pathway [22]. Some cytoplasmic or nuclear DNA receptors (cGAS, IFI16, AIM2, DAI, etc.) discovered in recent years are capable of specifically recognizing exogenous DNA. On the one hand, these DNA receptors induce the oligomerization of STING to induce the innate immune response through a series of signal transduction pathways [23,24,25]. On the other hand, similar to NLRs, they also induce the formation of inflammasomes and cause a strong inflammatory response [26,27]. Furthermore, it was reported that type III interferon (IFN-III) can also be produced in mucosal tissue cells through a similar pathway to IFN-I and demonstrate an intense antiviral activity [28,29,30,31]. A cytoplasmic DNA sensor ku70, which specifically induces IFN-III response, has been discovered recently [32]. After recognizing exogenous DNA, Ku70 translocates from the nucleus into the cytoplasm and induces the production of IFN-III through the STING-IRF1/7 signaling pathway, indicating a novel mechanism to initiate innate immune response [33,34,35].
Viral respiratory diseases have always been serious threats to human health. After infection, the patients usually display fever, sore throat, cough, and wheezing. In severe cases, such as HPAIV, SARS-CoV, and SARS-CoV-2 infections, patients usually display more severe systemic inflammatory responses or even succumb to death. With the outbreak of SARS-CoV-2 in late 2019, a growing number of studies have focused on viral respiratory diseases. This review is mainly focused on the influenza virus and coronavirus, two commonly observed respiratory viruses in terms of their intracellular RNA sensor-mediated innate immune responses following infection.
2. Influenza Virus
Influenza virus is a segmented single-stranded negative RNA virus of Orthomyxoviridae with high variability and transmissibility [36]. Segments of the viral genome encode 10 major proteins, among which two viral surface glycoproteins, hemagglutinin (HA) and neuraminidase (NA), are mainly used as the basis for viral typing [37,38,39,40]. Influenza virus is one of the most causative pathogens of human respiratory diseases, causing seasonal influenza and occasional influenza pandemics every year. Influenza viruses can be divided into four types: influenza A virus (IAV), influenza B virus (IBV), influenza C virus (ICV), and influenza D virus (IDV). Among them, IAV infects humans and a variety of animals, such as chickens, pigs, horses, marine mammals, etc, causing zoonotic diseases. H5N1, H7N9, and H1N1, the members of IAV that mainly infect poultry or mammals, pose a serious threat to public health. Considering that IAV is a segmented RNA virus, antigenic drift and antigenic shift may occur all the time during its replication due to the poor fidelity of RNA polymerase so that new subtypes of IAV could randomly and unexpectedly emerge. The new subtypes of IAV that better escape the host response would become dominant, and even break the interspecific barrier, resulting in cross-species transmission in humans [41]. Specifically, on the one hand, the IAV protein with accumulated amino acid mutations, especially NS1 [42] and PA-X [43], plays a major role in contributing to the evolution of IAV by inhibiting the host’s innate immune response through a variety of mechanisms: (i) Viral proteins directly interact with PRRs or innate immune molecules to inhibit the activation of the downstream signaling pathway (shown in Table 1) [44,45]; (ii) viral proteins bind to the host proteins [46,47] or dsDNA [48] to suppress the expression of antiviral genes; (iii) viral proteins bind to viral dsRNA to evade the innate immune recognition by RNA sensors [49]. On the other hand, the random reassortment of genome segments between different strains directly contributes to viral constant evolution and the generation of new subtypes [50,51]. It was reported that PB2 and PA gene segments containing mammalian adaptive mutations of H9N2 could be reassorted into the cocirculating H7N9 virus, resulting in the emergence of a new H7N9 virus genotype [52]. Thus, an in-depth understanding of the innate immune response caused by IAV RNA in different species would help to provide theoretical guidance for the control of IAV.
2.1. The Innate Immune Response to IAV Infection in Humans
In humans, viral RNA is primarily sensed by three types of PRRs, namely TLRs, RLRs, and NLRs [53,54,55]. Among them, TLRs are mainly responsible for the recognition of viral RNA in the endosome. Specifically, TLR3 senses dsRNA, while TLR7 and TLR8 sense single-stranded RNA (ssRNA) [56,57,58]. TLR9 recognizes CpG DNA or viral nucleic acids [59]. RLRs recognize dsDNA in the cytoplasm [60]. In addition, NLRs may also sense viral RNA, and then lead to the formation of inflammasomes [61,62].
It was reported that IAV replication intermediates could be recognized by TLR3 in human respiratory epithelial cells, which recruits TRIF and PI3K adapters and further activates IRFs and NF-κB signaling pathways [63,64]. In vivo, TLR3−/− mice displayed a higher viral titer and fewer inflammatory mediators upon H3N2 infection than wild-type (WT) mice, indicating the crucial role of TLR3 in resisting IAV infection [53,65]. However, despite the presence of TLR3, WT mice still show higher mortality, which may be attributable to their excessive inflammatory response [53]. Notably, the innate immune response mediated by TLR3 appears to be more inclined to induce the production of inflammatory cytokines rather than IFNs [66]. For IAV ssRNA, it can be sensed by TLR7 to activate NF-κB in a MyD88-dependent signaling pathway in pDCs, which contributes to the production of IFN-I [67,68]. Furthermore, the essential role of TLR7 in inflammatory cytokine activation was determined in TLR7−/− mice [69]. Although a few reports demonstrated that TLR8 was activated upon IAV infection [70,71], the mechanism by which TLR8 drives the innate immune response is slightly distinct from TLR7. TLR7 mainly induces the expression of Th17-polarizing cytokines, while TLR8 is closely related to the expression of Th1-type cytokines and IFN-I in CD14+ monocytes upon IAV infection [70], which suggests that TLR7 and TLR8 activate distinct pathways in monocytes. Moreover, TLR7 seems to play a role in the recognition of ssRNA to induce the IFN-I expression in dendritic cells, while TLR8 prefers to sense IAV ssRNA to induce the IL-12 expression in monocytes [71]. Thus, it is necessary to explore whether TLR7 or TLR8 has a cell-type preference upon IAV infection. It was found that the expression of TLR9 significantly increased upon H1N1 infection, suggesting that TLR9 may also be involved in the innate immune response against H1N1 infection [72].
In addition to TLRs, RLRs, as classical cytoplasmic RNA sensors, also play an important role in sensing viral RNA and combating viral infection [73,74]. It was reported that RIG-I was activated and engaged to MVAS on mitochondria upon IAV infection, initiating the expression of inflammatory cytokines and IFN-α/β in human alveolar epithelial cells [64,75]. It was also found that RIG-I could further enhance the expression of IFN-I through the MAVS-PI3K signaling pathway after recognizing the accumulated vRNA of IAV [76]. Furthermore, a recent study showed that RIG-I was present not only in the cytoplasm, but also in the nucleus [77]. The presence of RIG-I in the nucleus mainly senses vRNP in the process of IAV replication and induces the innate immune response, but the specific mechanism needs to be further investigated. However, it is still controversial whether MDA5, another representative member of RLR, contributes to the engagement of IAV dsRNA. Previous studies indicated that MDA5 was mainly involved in the innate immune response caused by some picornaviruses rather than IAV [78,79]. It was found that RIG-I acted as a primary PRR for IAV dsRNA in primary murine embryonic fibroblasts (MEFs), while MDA5 mainly functions as a transcriptional inducer, which benefits the amplification of ISG production [80]. In brief, MDA5 does not directly contribute to the engagement of IAV RNA, but it may participate in the amplification of host antiviral response by degrading viral nucleic acids through the OAS/RNase L system [80]. In addition, the length and structure of foreign RNA may also contribute to the differences in RNA recognition between RIG-I and MDA5. MDA5 mainly recognizes long dsRNA (>1 kb) and prefers to sense mRNA lacking ribose 2′-O methylation in the 5′cap structure, while RIG-I prefers to recognize short dsRNA sequences (<1 kb) [60,81,82]. Thus, more efforts will be required to investigate whether MDA5 is involved in IAV RNA recognition. Recently, it was found that IFI16, a well-known DNA receptor, also recognizes IAV RNA through the H1Na domain, eliciting an innate immune response by binding to RIG-I through another PYRIN domain to promote the activation of RIG-I [83]. It seems that our current understanding of nucleic acid receptors is quite limited, and there may be more nucleic acid sensors that can recognize DNA as well as RNA. In addition to IFN-I, there is no doubt that IFN-λ, a core molecule of innate mucosal immunity, is also abundantly expressed upon IAV infection [84,85,86,87]. However, the upstream signal transduction mechanisms of IFN-I and IFN-III are slightly different. IRFs play a dominant role in IFN-α/β expression, while NF-κB seems to be a key regulator of IFN-λ expression [88]. Furthermore, the IFN-β expression depends on the coordinated functions of a multifactor enhanceosome and the IFN-α expression on the IRF-binding cis-promoter elements, while the expression of IFN-III relies on the activation of NF-κB or IRFs alone, which suggests that there is a wider range of stimuli in inducing IFN-III expression [87,89,90]. These findings were further confirmed at the level of single-cell [91]. Although the sequence and structure between IFN-I receptors (IFNAR1/IFNAR2) and IFN-III receptors (IFNLR1/IL10R2) may vary, the downstream signal transduction process is highly similar, in which both trigger the expression of interferon stimulated gene (ISG) through the JAK-STAT signaling pathway [92,93].
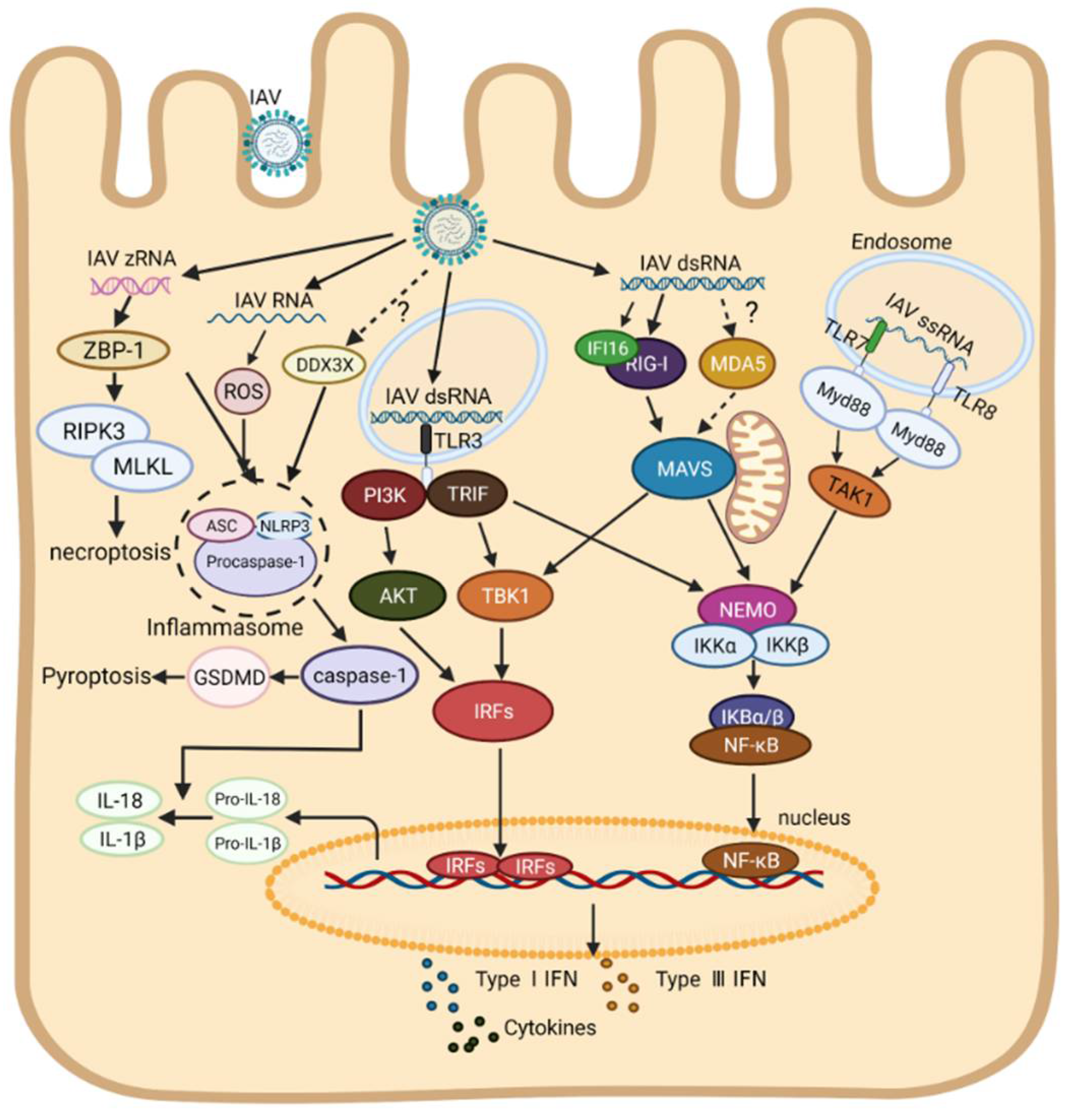
In recent years, the research on inflammasomes has become a hot area. It is now believed that two pathways concern the formation of inflammasomes, including the classical pathway mediated by caspase-1 and the non-classical pathway by caspase-11 [94,95,96,97]. It was reported that the expressions of NLRP3, caspase-1, pro-IL-1β, and pro-IL-18 were up-regulated via the NF-κB signaling pathway upon IAV infection, inducing the formation of inflammasomes in a caspase-1-dependent pathway [68]. IAV RNA or poly (I:C) can also be directly sensed by NLRP3 in human macrophages, resulting in pyroptosis and secreting a number of mature IL-1β and IL-18 [55,98,99]. Thus, it is very likely that NLRP3 acts as an intracellular RNA sensor for IAV to induce the formation of inflammasomes as well as inflammatory responses. Furthermore, a recent study indicates that DEAD-Box Helicase 3X (DDX3X), a member of the RNA helicase family protein, was also involved in the formation of NLRP3 inflammasome during WT IAV infection, but the specific ligands of DDX3X and related regulatory mechanisms remain unknown [100]. In addition to several known members in the caspase family (caspases-1, 4 and 11), more and more caspases have been shown to contribute to inflammasome formation. For instance, activated caspase-8 cleaves gasdermin D (GSDMD) during Yersinia infection, causing the formation of inflammasome [101,102], and activated caspase-3 can cleave GSDME, inducing pyroptosis and lung necrosis [103], and activated caspase-6 promotes inflammasome formation though activating the ZBP1-RIPK3 complex and induces a strong inflammatory response upon IAV infection [104]. ZBP-1, known as an intracellular dsDNA sensor, was reported to regulate NLRP3 inflammasome activation upon IAV infection [105]. Recently, it was found that ZBP-1 could sense IAV Z-RNA, which promotes the activation of mixed lineage kinase domain-like pseudokinase (MLKL) mediated by RIPK-3, resulting in necroptosis [106]. It seems that ZBP-1 may serve as a new PRR for IAV RNA inhost, which furthered our understanding of the mechanism of cell death and innate immune response caused by IAV RNA. As shown in Figure 1, the RNA sensors-mediated innate immune signal transduction pathways to IAV infection in human cells are summarized (Figure 1).
2.2. The Innate Immune Response to IAV Infection in Chickens
Since the avian is one of the main hosts of IAV infection, the IAV from the avian is usually described as the avian influenza virus (avian IAV). Avian IAV is divided into HPAIV and low pathogenic avian influenza virus (LPAIV) according to its pathogenicity. HPAIVs mainly consist of H5 or H7 subtypes [107], not only causing mortality in chickens, but also posing a serious threat to public health [108]. Similar to humans, avian IAV RNA is primarily sensed by TLR3 and TLR7 [109,110,111]. It was reported that the TLR3 and IFN-β mRNA levels were significantly upregulated in the brain, spleen, and lungs in H5N1-infected chickens [112]. Meanwhile, the same change was observed under the treatment by poly (I:C) in chickens [112,113,114]. Furthermore, homologs of several key proteins in mammalian TLR3 signal transduction pathway were also been identified in chickens, such as JNK, TRIF, TBK1, IκκE, etc. [111,115]. Thus, chicken TLR3 (chTLR3) may act as the avian IAV RNA sensor in chickens, performing similar functions as mammalian TLR3. However, the TLR3 signaling cascade in chickens has not been systematically characterized. Due to the gene sequence of chTLR8 was highly disrupted in chickens [116], instead, chTLR7 may play a major role in the recognition of avian IAV ssRNA in endosomes [116,117]. It was found that the expression of IL-Iβ is highly upregulated upon the stimulation of synthetic ssRNA, a chTLR7 ligand, leading to the antiviral response in vitro [118]. It was also found that the mRNA level of IL-Iβ was significantly upregulated by the treatment of chTLR7 agonist, while the IFN-I mRNA level had no change [116], which suggests that chTLR7 prefers to induce the production of proinflammatory cytokines rather than IFN-I. Since RIG-I is congenitally deficient in chickens [119], thus dsRNA in the cytoplasm is mainly recognized by chMDA5, which may account for the high susceptibility of chickens to avian IAV infection [120,121,122]. It was reported that avian IAV RNA was mainly sensed by chMDA5, which further activates IRF7 and NF-κB signaling pathways through interacting with chSTING, initiating IFN response [123,124]. Of note, due to the natural absence of IRF3 [111], the innate immune response is mainly mediated by IRF1 and/or IRF7 in chickens [125,126,127]. In addition, chicken DDX3X (chDDX3X) could also induce IFN-β production through the chDDX3X-chSTING-chIRF7 signaling pathway [128]. Thus, it seems that the RLRs signal transduction in chickens is highly dependent on chMDA5 and chSTING. Unfortunately, few reports are currently available regarding the formation of inflammasome induced by avian IAV RNA in chickens. More efforts will be required to investigate avian IAV-induced pyroptosis in the future.
2.3. The Innate Immune Response to IAV Infection in Other Species
In addition to humans and chickens, IAV also replicates in other species. In comparison with chickens, the homolog of RIG-I was present in ducks, which mediates innate antiviral responses through its CARD domain [129,130]. It was reported that RIG-I and MDA5 were all involved in the recognition of avian IAV RNA in ducks and then elicited the innate immune response through NF-κB or IRF7 signaling pathways [131,132,133]. In addition, TLR7 and TLR3 were also significantly activated upon avian IAV infection [134]. It seems that the RNA sensors in ducks are more complete than that of chickens, which may explain the higher resistance of ducks to avian IAV infection than chickens. Swine, another host for IAV, play an important role in IAV transmission and mutation, which results in a risk of bidirectional infection between humans and swine [135]. In particular, the outbreaks of the H1N1, a mutant strain in swine, have posed a severe threat to human health [136]. Thus, it is necessary to evaluate the innate immune response caused by IAV in swine. It was found that the expressions of TLR3, TLR7 and RIG-I in porcine alveolar macrophages were up-regulated during IAV infection, and the JAK-STAT and MAPK signaling pathways were highly activated [137,138]. However, MDA5 seems to contribute little to the recognition of IAV RNA in swine [139].
2.4. Regulation of the Innate Immune Response by MiRNAs and Viral Proteins upon IAV Infection
It was found that varied miRNAs were involved in the innate immune response upon IAV infection in humans [140]. On the one hand, some miRNAs play a positive role, inducing an antiviral immune response and inhibiting IAV replication. For example, the expression of miR-93 in alveolar epithelial cells infected with IAV could be significantly down-regulated, resulting in the release of the target protein JAK, which mediated innate immune response through the JAK-STAT signaling pathway [141]. The expression of miR-340 was down-regulated in IAV-infected A549 cells, relieving the inhibition of RIG-I and OAS2 and enhancing the innate immune response [142]. On the other hand, the other miRNAs play a negative role in innate immune response upon IAV infection. For instance, miR-146a-5p is up-regulated in human nasal epithelial cells (HNECs) infected with H3N2, and negatively regulates TRAF6 expression [143]. miR-29c enhances the expression of A20 protein induced by IAV, inhibiting the activity of NF-κB and the expression of pro-inflammatory cytokines [144]. Previous reviews have well documented the multiple roles of miRNAs in the regulation of innate immunity upon IAV infection [140,145]. In addition to miRNAs, other noncoding RNAs such as long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs) may also involve regulation of mRNAs in control of gene transcription. Furthermore, some IAV viral proteins, as listed in Table 1, were also found directly suppress the innate immune response in different species.

{kind=link}
{kind=link}
{kind=link}
Table 1.
The roles of IAV proteins in innate immune response across the species.
| Host | Proteins | Protein Function | References |
|---|---|---|---|
| Human | NS1 | Inhibit the activation of RIG-I | [146] |
| Inhibit the ubiquitination of RIG | [147,148,149] | ||
| Inhibit the formation of NLRP3 inflammasome | [150] | ||
| Bind with IKK and Inhibit the activation of NF-κB | [151,152] | ||
| Inhibit the ubiquitination of TRAF3 | [153] | ||
| Degradation of sphingosine 1-phosphate lyase (SPL) and suppress IKKϵ-mediated type I IFN response | [154] | ||
| PB1-F2 | Destroy the mitochondrial membrane | [155,156] | |
| Impair innate immunity by inducing mitophagy | [157] | ||
| Interact with IKKβ and Inhibit the activation of NF-κB | [158] | ||
| Interact with IRF3 and reduced the expression of IFN-β | [159] | ||
| PB1 | Degrade the MAVS by autophagy | [160] | |
| PA-X | Degrade viral dsRNA | [161] | |
| Inhibit the RIG-I-MAVS signaling pathway | [162] | ||
| Inhibit Ankrd17-mediated immune response | [163] | ||
| inhibit NF-κB transcription | [164] | ||
| Chicken | NS1 | Act in concert with polymerase complexes | [165] |
| PB1-F2 | Interact with MAVS and inhibit the IFN response | [166] | |
| Duck | NS1 | Inhibit the MDA5-mediated signaling pathway | [133] |
| PB1-F2 | Inhibit RIG-I ubiquitination | [167] | |
| Swine | NS1 | Impair ASC speck formation and inhibit IL-1β production | [168] |
3. Coronavirus
Coronavirus is a linear single-stranded RNA virus, belonging to the genus of Corona-virus in the family Coronaviridae [169]. According to the phylogeny, it can be classified into four genera: α, β, γ, and δ [170]. Coronavirus can infect a variety of mammals and poultry, including humans. At present, severe respiratory symptoms are consistent in humans and poultry, while distinct gastrointestinal symptoms are present in swine [171]. Therefore, it remains a high demand to explore the innate immune response upon coronavirus infection. The innate immune response to coronavirus infection in different species is reviewed as follows.
3.1. The Innate Immune Response to Coronavirus Infection in Humans
Up to now, three types coronavirus pose a severe threat to public health, namely SARS-CoV, SARS-CoV-2 and MERS-CoV, all of which belong to β-Coronavirus [170,172,173]. Both SARS-CoV and SARS-CoV-2 have caused global pandemics, especially SARS-CoV-2, which has caused millions of people deaths and uncountable economic losses worldwide since its outbreak in late 2019 [169,174]. With the constant evolution of SARS-CoV-2, a variety of evolved strains such as the Delta mutant strain [175] and Omicron mutant strain [176] have emerged, which results in enormous challenges for its prevention and control. Previous studies reported that SARS-CoV inhibited the activation of IFN-I promoter in the early stage after infection [177,178], and SARS-CoV-2 showed a similar effect [179]. The early immunosuppression in host could benefit the proliferation of the coronavirus, leading to delayed production of inflammatory cytokines and causing serious clinical symptoms [177,180].
Similar to IAV, the RNA sensors for coronavirus are TLRs and RLRs [181,182]. In vivo, TLR3−/− or TRIF−/− mice showed an increased susceptibility to SARS-CoV infection compared to that of wild-type (WT) mice, displaying a more severe lesion in the lung and a higher viral titer in the serum than the WT mice [183]. It was, not until recently, found that TLR3 and TLR7, as direct intracellular RNA sensors, were highly activated in SARS-CoV-2-infected CalU-3/MRC-5 multicellular spheroids in the early stage of infection. Particularly, TLR3 activates the expression of IL-1β, IL-4, IL-6 and IFN-I largely through the IRF3 signaling pathway within 48 h after SARS-CoV-2 infection, while IRF7 seems to induce the expression of IFN-I and IFN-III through the activation of NF-κB after 48 h of viral infection [184]. These findings provide direct evidence that TLR3 acts as an RNA sensor of coronavirus. In addition, it was reported that severe cases with SARS-CoV-2 infection have more multiple mutations in TLR3 than those with mild clinical symptoms, and these mutations are closely related to the severity of clinical symptoms, suggesting that the mutations of TLR3 may be the potential reason for severe clinical symptoms [185]. Furthermore, it was found that the polymorphism of L412F in TLR3 was more closely related to the severity of SARS-CoV-2 infection [186]. These findings indicate a critical role of TLR3 in recognizing the coronavirus RNA, triggering an innate immune response to SARS-CoV-2 infection.
It was found that TLR7/8 played an indispensable role in the recognition of corona-virus ssRNA, triggering host response [187,188]. SARS-CoV ssRNA could be engaged by TLR7 in plasmacytoid dendritic cells (pDCs), eliciting a rapid innate immune response to suppress viral replication [189]. Furthermore, it was also found that TLR7 functioned as a primary sensor for viral ssRNA upon MERS-CoV infection, modulating the IFN-I response [190]. In early 2020, SARS-CoV-2 broke out worldwide [174]. In order to better understand the innate immune response caused by SARS-CoV-2, bioinformatics analysis was used to identify the ssRNA sequences sensed by TLR7/8 in the whole genomes of SARS-CoV-2, SARS-CoV and MERS-CoV. It was found that a number of viral ssRNA fragments could be sensed by TLR7/8, which provides coronavirus with a shortcut to trigger an innate immune response [191]. It should be noted that more UU (U/C) and UU (G/A) motifs existed in SARS-CoV-2 genomes compared with that of SARS-CoV, which may lead to increased inflammatory response and aggravated severe clinical symptoms [187]. Clinically, the patients, especially males, with TLR7 loss of function variants showed higher susceptibility to SARS-CoV-2 and lower expression level of IFN-I and IFN-II [192], while mutations in TLR8 seems to have no effect on its receptor function [193]. Taken together, lines of evidence show that both TLR7 and TLR8 play an essential role in sensing coronavirus ssRNA to initiate the innate immune response, and the mutations of TLR7 may be used as an indicator to evaluate the human susceptibility to SARS-CoV-2. Of note, since the genes encoding TLR7 and TLR8 are present on the X chromosome, sex preference occurred upon coronavirus infection, especially SARS-CoV-2, which is manifested by the low expression level of IL-6 and relatively mild clinical symptoms in women [194,195].
Different from IAV, coronavirus genome in the cytoplasm of host cells is primarily sensed by MDA5 and LGP2, consequently activating innate immune response [196,197,198]. It was found that SARS-CoV-2 replication intermediates specifically activated a delayed IFN-I response through MDA5, LGP2 and NOD1 in CalU 3 cells, while RIG-I did not affect IFN-I response [196]. Surprisingly, NOD1, a well-known sensor for bacterial peptidoglycans, was identified as a PRR sensing SARS-CoV-2 [196]. It would be interesting to explore the mechanism by which NOD1 triggers innate immune response upon SARS-CoV-2 infection. As for RIG-I, another important dsRNA sensor in the cytoplasm, its role in the innate immune response during SARS-CoV-2 infection remains controversial. It was early found that RIG-I had no contribution to IFN-β production upon SARS-CoV-2 infection [196,199], while a recent study indicated that the IFN-β expression significantly decreased in RIG-I−/− CalU 3 cells [200]. Furthermore, it was reported that the RIG-I helicase domain (HD) was able to directly interact with the 3′UTR of SARS-CoV-2 RNA in human alveolar epithelial cells, and inhibited viral replication by blocking the binding of RNA-dependent RNA polymerase (RdRp) to viral RNA [201], suggesting that RIG-I plays a role in the host response to SARS-CoV-2 infection. Thus, it seems that RIG-I inhibits viral replication in an IFN-independent manner upon SARS-CoV-2 infection.
Evidence has shown that severe forms of SARS-CoV or SARS-CoV-2 infections were closely related to acute severe inflammatory reactions [202,203]. Multiple proinflammatory cytokines and inflammasome derivatives, such as IL-1β, IL-18, NLRP3 and LDH, were detected in the serum of patients infected with SARS-CoV-2, which suggests that a strong inflammatory response caused by SARS-CoV-2 may be closely related to inflammasome formation [203,204,205]. Persistent activation of NLRP3 inflammasomes can lead to severe clinical symptoms, including fever, necrosis, and severe inflammatory responses [206]. A recent study analyzed the inflammasome activation in the serum of 129 SARS-CoV-2 patients, and found that the expression of caspase-1 p20 and IL-18 in the serum was significantly elevated [207]. Furthermore, the NLRP3-ASC complex was observed in peripheral blood mononuclear cells (PBMCs), which indicates that NLRP3 is highly activated upon SARS-CoV-2 infection [207]. Moreover, a considerable amount of cleaved GSDMD, an indicator of pyroptosis, was also observed using multiplex immunohistochemistry [208]. These results indicate that SARS-CoV-2 is capable of initiating NLRP3 activation. Nevertheless, the effect of coronavirus RNA on the formation of inflammasomes is still not clear. It was, not until recently, found that GU-rich single-stranded RNA (GU-rich RNA) derived from SARS-CoV or SARS-CoV-2 could initiate a TLR8-dependent pro-inflammatory response in human macrophages without pyroptosis. GU-rich RNA can be engaged by TLR8 and activates inflammasomes through the TLR8-Caspase 8-RIPK3-NLRP3 signaling pathway, and releases mature IL-1β, IL-6 and TNF, which provides preliminary data favoring the involvement of coronavirus RNA in inflammasome formation [209,210] Further studies are required to determine whether coronavirus RNA triggers inflammatory responses through the classical pyroptotic pathway. Given the current pandemic of SARS-CoV-2, to better understand the innate immune recognition of coronaviruses with cross-species transmission capability, we comprehensively compared the innate sensing of three coronaviruses, MERS-CoV, SARS-CoV, and SARS-CoV-2 (Table 2).
Interestingly, it was found that the cGAS-STING signaling pathway, a signaling pathway activated by recognition of DNA by cellular cGAS, was also activated to drive a robust IFN-I response after SARS-CoV-2 infection [218,219]. Different from the direct sense for viral RNA by TLRs and RLRs, cGAS-STING activation mainly relies on the recognition of DNA from either pathogens or host tissue damages [220]. RNA sensors-mediated signal transduction pathways in the innate immune response to coronavirus infection in humans are summarized and shown in Figure 3.
3.2. The Innate Immune Response to Coronavirus Infection in Chickens
Avian infectious bronchitis virus (IBV), a representative avian coronavirus, mainly causes respiratory symptoms, but the lack of available cell lines for viral infection has limited its further investigation [221]. Thus, the current research on IBV is mainly focused on vivo experiments. At present, due to the emergence of mutant strains, the control of IBV is becoming more and more difficult [222]. An in-depth understanding of the innate immune response to IBV infection is very important. Existing evidence suggests that chTLR3, chTLR7, and chMDA5 are involved in the recognition of IBV RNA to trigger the innate immune response [223]. Upon IBV M41 strain infection, the TLR3-TRIF signaling pathway was highly activated, which further triggered the innate immune response and inhibited IBV replication [224,225]. In contrast, it was found that the mRNA level of chTLR3 was down-regulated in respiratory epithelial cells early during IBV Connecticut strain infection [226]. Furthermore, the effect of IBV infection on the chTLR7 signaling pathway is still not very clear [227]. It was reported that the expression of chMDA5 and chSTING were up-regulated in IBV-infected birds, which further initiated the innate immune response [225,228]. chLGP2, a member of RLRs with less investigated in chicken, was reported to interact with chTRBP to dampen IBV infection [229]. The research on the innate immune response to coronavirus infection in chickens is quite limited and should be highly encouraged.
3.3. The Innate Immune Response to Coronavirus Infection in Other Species
Porcine coronavirus consists of porcine respiratory coronavirus (PRCV), porcine delta coronavirus (PDCoV), porcine epidemic diarrhea virus (PEDV), transmissible gastroenteritis virus (TGEV), etc. It was reported that viral RNA could be sensed by TLR3 and TLR9 to activate the NF-κB signaling pathway in PEDV-infected IECs, which is independent of RIG-I [230]. Meanwhile, the mRNA levels of MDA5 were significantly up-regulated in TGEV-infected porcine kidney cells, promoting the expression of inflammatory cytokines [231]. Of note, a recent study found that PDCoV infection robustly activated the expression of RIG-I, thus enhancing the IFN-β production [232]. It was also demonstrated that TLR7 expression was significantly upregulated and elicited IFN response after PEDV infection in IPEC-J2 cells [233]. Furthermore, it was recently found that NLRP3 was highly activated in porcine coronavirus infected cells, leading to pyroptosis as demonstrated by cleaved GSDMD and IL-1β release [234], suggesting that NLRs may play an important role in the innate immune response to porcine coronavirus infection.
3.4. The Innate Immune Regulation by MiRNAs and Viral Proteins during Coronavirus Infection
Up to now, there are few reports regarding the regulation of innate immune response by miRNAs during coronavirus infection in humans. It was found that miRNAs were involved in innate immune response, viral binding, viral invasion, and intracellular stress after coronavirus infection [235,236,237,238]. Angiotensin-converting enzyme 2 (ACE2), which serves as a receptor for coronavirus, can interact with transmembrane serine protease 2 (TMPRSS2) to facilitate viral entry [239]. It was reported that miR-200c could target the 3′-UTR of ACE2 mRNA, reducing the ACE expression and suppressing viral entry [240]. Similar to miR-200c, miR-98-5p suppressed the expression of TMPRSS2 [241]. Interestingly, a recent study identified four unique microRNA-like small RNAs encoded by SARS-CoV-2, namely SCV2-miR-1ab-1-3p, SCV2-miR-1ab-2-5p, SCV2-miR-1ab-3-5p, and SCV2-miR-3a-5p, which targets the gene of IFN-I signaling pathway, regulating innate immunity [242]. Furthermore, two virus-derived miRNA isoforms were identified in SARS-CoV-2-infected human cells, namely CoV-2-miR-O7a.1 and CoV-2-miR-O7a.2. CoV-2-miR-O7a.2 suppresses ISGs expression by targeting its 3′UTR and evades the IFN-mediated immune response [243]. Except for the limited research, current research performed a functional enrichment analysis regarding the roles of miRNAs in the inflammatory response in SARS-CoV-2-infected patients. Specifically, three miRNAs were significantly down-regulated in SARS-CoV-2-infected patients, namely miR-26a-5p, miR-29b-3p, and miR-34a-5p. Among them, miR-26a-5p may regulate inflammatory response by targeting IL-6, and miR-34a-5p is predicted to regulate inflammation by targeting Caspase-1 [244].
In contrast to miRNAs, the roles of coronavirus proteins in the innate immune response have been widely investigated across the species (Table 3).
4. The Differences in Innate Immune Response between Species during IAV/Coronavirus Infections
Innate immune response is highly different between species for both IAV and coronavirus infections. On the one hand, the differences in PRRs between species directly influence the host’s innate immune recognition. For instance, the congenital deficiency of RIG-I in chicken enhances its susceptibility to these two viruses greatly [119]. On the other hand, the discrepancy in protein-protein interactions (PPIs) modulated by innate immune recognition of viral RNA also contributes to the differences in innate immune responses across the species. It was reported that the MAVS-IRF3 signaling pathway plays an important role in innate immune response in humans [292], whereas in chickens, it seems to be more inclined to induce innate immune response via the STING-IRF7/MAVS-IRF7 signaling pathway [128]. Damaged dsDNA could be recognized by TLR9/cGAS upon coronavirus infection, thereby inducing an innate immune response via the TLR9-MyD88/cGAS-STING signaling pathway in humans [220,293], but similar mechanism has not been identified in chickens. In addition, it was found that NOD1, an RNA sensor for corona-virus [196], could recognize poly (I:C) dsRNA and induce an innate immune response by binding to MDA5 in humans [294], whereas no similar report has been found in other species. Furthermore, a recent study found that PDCoV infection induced the expression of IFN-β via the RIG-MAVS signaling pathway in swine [232], however, RIG-1 initiates host response in an IFN-independent way by competitively binding to the coronavirus RNA with RdRp in humans [201].
5. Conclusions
Up to now, IAV and coronavirus have posed a serious threat to animals and public health. As the first line of host defense against virus infection, innate immune response is particularly important. TLRs, NLRs, and RLRs play pivotal roles in innate immune response via recognizing IAV and coronavirus RNAs to trigger innate immune signaling pathways. Both IAV and coronavirus RNAs in endosomes are sensed by TLR3 and TLR7. However, IAV appears to trigger the innate immune response mainly through RIG-I, whereas coronavirus prefers to rely on MDA5. It is still unclear whether there are any RNA sensors currently unknown to us and are involved in the innate immune response to IAV and coronavirus infections. However, the innate immune recognition of IAV/coronavirus RNA also varies considerably across species, including the differences in PRRs and PPIs modulated by innate immune recognition of viral RNA. This review primarily focuses on PRRs-mediated innate immune signaling and provides an overview of the roles of RNA sensors in host innate response to IAV and coronavirus infections in different species. A comprehensive understanding of the mechanisms by which these two respiratory viral pathogens initiate the innate immune response via RNA sensors will help us better understand the difference in susceptibility across the species and develop novel vaccines or antiviral drugs for their effective control.
Author Contributions
Conceived and designed: S.J.Z.; wrote the paper: W.L. and H.W.; revised the paper: S.J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (# 32130105) and the earmarked fund for CARS-40, China.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role.
References
- Uematsu, S.; Akira, S. Toll-like receptor and innate immunity. Seikagaku 2007, 79, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Tseng, J.C.; Yang, J.X.; Liu, Y.L.; Yeh, D.W.; Lai, C.Y.; Yu, G.Y.; Hsu, L.C.; Huang, C.M.; Chuang, T.H. Toll-Like Receptor 21 of Chicken and Duck Recognize a Broad Array of Immunostimulatory CpG-oligodeoxynucleotide Sequences. Vaccines 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Ruemmele, F.M.; Beaulieu, J.F.; Lentze, M.J. LPS-signaling in intestinal epithelial cells requires a functional Toll-like receptor 4. Gastroenterology 2002, 122, A152. [Google Scholar]
- Akira, S.; Hoshino, K.; Kaisho, T. The role of Toll-like receptors and MyD88 in innate immune responses. J. Endotoxin Res. 2000, 6, 383–387. [Google Scholar] [CrossRef]
- Wesche, H.; Henzel, W.J.; Shillinglaw, W.; Li, S.; Cao, Z.D. MyD88: An adapter that recruits IRAK to the IL-1 receptor complex. Immunity 1997, 7, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Fujita, T. RIG-I family RNA helicases: Cytoplasmic sensor for antiviral innate immunity. Cytokine Growth Factor Rev. 2007, 18, 545–551. [Google Scholar] [CrossRef]
- Yoneyama, M.; Fujita, T. Function of RIG-I-like receptors in antiviral innate immunity. J. Biol. Chem. 2007, 282, 15315–15318. [Google Scholar] [CrossRef] [Green Version]
- Bowie, A.G.; Fitzgerald, K.A. RIG-I: Tri-ing to discriminate between self and non-self RNA. Trends Immunol. 2007, 28, 147–150. [Google Scholar] [CrossRef]
- Newbrough, S.A.; Saito, T.; Gale, M. Characterizing the Role of LGP2 in RLR Signaling and AntiViral Immunity. J. Allergy Clin. Immun. 2011, 127, Ab91. [Google Scholar] [CrossRef]
- Loo, Y.M.; Fornek, J.; Crochet, N.; Zeng, H.; Akira, S.; Gill, M.A.; Tumpey, T.M.; Garcia-Sastre, A.; Katze, M.G.; Gale, M. Distinct RIG-I and MDA5 signaling regulation by RNA viruses in innate immunity. J. Interf. Cytok Res. 2007, 27, 697. [Google Scholar]
- Seth, R.B.; Sun, L.J.; Ea, C.K.; Chen, Z.J.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappa B and IRF3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trindade, B.C.; Chen, G.C.Y. NOD1 and NOD2 in inflammatory and infectious diseases. Immunol. Rev. 2020, 297, 139–161. [Google Scholar] [CrossRef]
- Le Bourhis, L.; Benko, S.; Girardin, S.E. Nod1 and Nod2 in innate immunity and human inflammatory disorders. Biochem. Soc. Trans. 2007, 35, 1479–1484. [Google Scholar] [CrossRef] [Green Version]
- Ruefli-Brasse, A.A.; Lee, W.P.; Hurst, S.; Dixit, V.M. Rip2 participates in Bcl10 signaling and T-cell receptor-mediated NF-kappa B activation. J. Biol. Chem. 2004, 279, 1570–1574. [Google Scholar] [CrossRef] [Green Version]
- Franchi, L.; Eigenbrod, T.; Munoz-Planillo, R.; Nunez, G. The inflammasome: A caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat. Immunol. 2009, 10, 241–247. [Google Scholar] [CrossRef]
- Martinon, F.; Mayor, A.; Tschopp, J. The Inflammasomes: Guardians of the Body. Annu. Rev. Immunol. 2009, 27, 229–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.J.; Zhao, Y.; Wang, K.; Shi, X.Y.; Wang, Y.; Huang, H.W.; Zhuang, Y.H.; Cai, T.; Wang, F.C.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Underhill, D.M.; Rossnagle, E.; Lowell, C.A.; Simmons, R.M. Dectin-1 activates Syk tyrosine kinase in a dynamic subset of macrophages for reactive oxygen production. Blood 2005, 106, 2543–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thome, M. Multifunctional roles for MALT1 in T-cell activation. Nat. Rev. Immunol. 2008, 8, 495–500. [Google Scholar] [CrossRef]
- Unterholzner, L.; Keating, S.E.; Baran, M.; Horan, K.A.; Jensen, S.B.; Sharma, S.; Sirois, C.M.; Jin, T.C.; Latz, E.; Xiao, T.S.; et al. IFI16 is an innate immune sensor for intracellular DNA. Nat. Immunol. 2010, 11, 997–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Long, Z.; Jia, H.; Chen, G.; Fang, Y.; Wang, G.; Jing, Z. PRV infection triggers innate antiviral immune responses via cGAS-STING axis and TLR9-MyD88 axis mediated DNA-sensing signaling pathway. Eur. J. Immunol. 2019, 49, 1078–1079. [Google Scholar]
- Takaoka, A.; Wang, Z.; Choi, M.K.; Yanai, H.; Negishi, H.; Ban, T.; Lu, Y.; Miyagishi, M.; Kodama, T.; Honda, K.; et al. DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 2007, 448, 501–505. [Google Scholar] [CrossRef]
- Fernandes-Alnemri, T.; Yu, J.W.; Datta, P.; Wu, J.; Alnemri, E.S. AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 2009, 458, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Hornung, V.; Ablasser, A.; Charrel-Dennis, M.; Bauernfeind, F.; Horvath, G.; Caffrey, D.R.; Latz, E.; Fitzgerald, K.A. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 2009, 458, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Ank, N.; Paludan, S.R. Type III IFNs: New layers of complexity in innate antiviral immunity. Biofactors 2009, 35, 82–87. [Google Scholar] [CrossRef]
- Syedbasha, M.; Egli, A. Interferon Lambda: Modulating Immunity in Infectious Diseases. Front. Immunol. 2017, 8, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, R.P.; Kotenko, S.V. Interferon-lambda: A new addition to an old family. J. Interferon Cytokine Res. 2010, 30, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, P.; Kindsvogel, W.; Xu, W.F.; Henderson, K.; Schlutsmeyer, S.; Whitmore, T.E.; Kuestner, R.; Garrigues, U.; Birks, C.; Roraback, J.; et al. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nat. Immunol. 2003, 4, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Brann, T.W.; Lidie, K.B.; Zhou, M.; Oguariri, R.M.; Yang, J.; Fullmer, B.; Zheng, X.; Degray, G.S.; Imamichi, H.; et al. Ku70 is a novel cytosolic DNA sensor and induces type-III IFN rather than Type-I IFN. Cytokine 2010, 52, 42–43. [Google Scholar] [CrossRef]
- Imamichi, T.; Zhang, X.; Zhou, M.; Lempicki, R.; Baseler, M.; Veenstra, T.; Young, H.; Lane, H.C. Ku70 is a novel cytosolic DNA sensor that induces a Type-III rather than Type-I IFN via activation of IRF-1 and IRF-7. J. Immunol. 2011, 186, 4541–4545. [Google Scholar]
- Sui, H.Y.; Zhou, M.; Imamichi, H.; Jiao, X.L.; Sherman, B.T.; Lane, H.C.; Imamichi, T. STING is an essential mediator of the Ku70-mediated production of IFN-gimel 1 in response to exogenous DNA. Sci. Signal. 2017, 10, eaah5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, H.Y.; Chen, Q.; Imamichi, T. Cytoplasmic-translocated Ku70 senses intracellular DNA and mediates interferon-lambda1 induction. Immunology 2021, 163, 323–337. [Google Scholar] [CrossRef]
- Fujiyoshi, Y.; Kume, N.P.; Sakata, K.; Sato, S.B. Fine-Structure of Influenza-a Virus Observed by Electron Cryomicroscopy. EMBO J. 1994, 13, 318–326. [Google Scholar] [CrossRef]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and Ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Engelhardt, O.G.; Fodor, E. Functional association between viral and cellular transcription during influenza virus infection. Rev. Med. Virol. 2006, 16, 329–345. [Google Scholar] [CrossRef]
- Yewdell, J.W.; Ince, W.L. Frameshifting to PA-X Influenza. Science 2012, 337, 164–165. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.R. The PB1-F2 protein of Influenza A virus: Increasing pathogenicity by disrupting alveolar macrophages. Virol. J. 2007, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Ciminski, K.; Chase, G.P.; Beer, M.; Schwemmle, M. Influenza A Viruses: Understanding Human Host Determinants. Trends Mol. Med. 2021, 27, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Nogales, A.; Martinez-Sobrido, L.; Topham, D.J.; DeDiego, M.L. Modulation of Innate Immune Responses by the Influenza A NS1 and PA-X Proteins. Viruses 2018, 10, 708. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Ma, C.; Liu, X. PA-X: A key regulator of influenza A virus pathogenicity and host immune responses. Med. Microbiol. Immunol. 2018, 207, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Marc, D. Influenza virus non-structural protein NS1: Interferon antagonism and beyond. J. Gen. Virol. 2014, 95, 2594–2611. [Google Scholar] [CrossRef] [PubMed]
- Klemm, C.; Boergeling, Y.; Ludwig, S.; Ehrhardt, C. Immunomodulatory Nonstructural Proteins of Influenza A Viruses. Trends Microbiol. 2018, 26, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Noah, D.L.; Twu, K.Y.; Krug, R.M. Cellular antiviral responses against influenza A virus are countered at the posttranscriptional level by the viral NS1A protein via its binding to a cellular protein required for the 3’ end processing of cellular pre-mRNAS. Virology 2003, 307, 386–395. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Krug, R.M. Influenza A virus NS1 protein targets poly(A)-binding protein II of the cellular 3’-end processing machinery. EMBO J. 1999, 18, 2273–2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasina, M.; Le May, N.; Bugai, A.; Fu, Y.; Soderholm, S.; Gaelings, L.; Ohman, T.; Tynell, J.; Kyttanen, S.; Barboric, M.; et al. Influenza virus NS1 protein binds cellular DNA to block transcription of antiviral genes. Biochim. Biophys. Acta 2016, 1859, 1440–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornholdt, Z.A.; Prasad, B.V. X-ray structure of NS1 from a highly pathogenic H5N1 influenza virus. Nature 2008, 456, 985–988. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Wang, S.G.; Yin, Y.B.; Zhang, G.Z.; Carter, R.A.; Wang, J.L.; Xu, G.L.; Sun, H.L.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc. Natl. Acad. Sci. USA 2015, 112, 548–553. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.Y.; Ke, C.W.; Li, Q.; Yuan, R.Y.; Xiang, D.; Jia, W.X.; Yu, Y.D.; Liu, L.; Huang, C.; Qi, W.B.; et al. Novel Reassortant Avian Influenza A(H5N6) Viruses in Humans, Guangdong, China, 2015. Emerg. Infect. Dis. 2016, 22, 1507–1509. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Yin, Y.; Liu, J.; Wang, X.; Zhou, Y.; Wang, Z.; Sun, Y.; Sun, H.; Li, F.; Song, J.; et al. Reassortment with dominant chicken H9N2 influenza virus contributed to the fifth H7N9 virus human epidemic. J. Virol. 2021, 95, e01578-20. [Google Scholar] [CrossRef] [PubMed]
- Le Goffic, R.; Balloy, V.; Lagranderie, M.; Alexopoulou, L.; Escriou, N.; Flavell, R.; Chignard, M.; Si-Tahar, M. Detrimental contribution of the Toll-like receptor (TLR)3 to influenza A virus-induced acute pneumonia. PLoS Pathog. 2006, 2, e53. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P.Y. The NLRP3 Inflammasome Mediates In Vivo Innate Immunity to Influenza A Virus through Recognition of Viral RNA. Immunity 2009, 30, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Funami, K.; Tanabe, M.; Oshiumi, H.; Shingai, M.; Seto, Y.; Yamamoto, A.; Seya, T. Subcellular localization of Toll-like receptor 3 in human dendritic cells. J. Immunol. 2003, 171, 3154–3162. [Google Scholar] [CrossRef] [Green Version]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- Boehme, K.W.; Compton, T. Innate sensing of viruses by toll-like receptors. J. Virol. 2004, 78, 7867–7873. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Bauernfried, S.; Scherr, M.J.; Pichlmair, A.; Duderstadt, K.E.; Hornung, V. Human NLRP1 is a sensor for double-stranded RNA. Science 2021, 371, eabd0811. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Levine, S.J. Toll-like receptor 3, RIG-I-like receptors and the NLRP3 inflammasome: Key modulators of innate immune responses to double-stranded RNA viruses. Cytokine Growth Factor Rev. 2011, 22, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Guillot, L.; Le Goffic, R.; Bloch, S.; Escriou, N.; Akira, S.; Chignard, M.; Si-Tahar, M. Involvement of toll-like receptor 3 in the immune response of lung evithelial cells to double-stranded RNA and influenza A virus. J. Biol. Chem. 2005, 280, 5571–5580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.X.; Zhang, W.; Duggan, E.S.; Booth, J.L.; Zou, M.H.; Metcalf, J.P. RIG-I and TLR3 are both required for maximum interferon induction by influenza virus in human lung alveolar epithelial cells. Virology 2015, 482, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, C.Y.; Jin, Y.; Zou, S.M.; Qi, P.; Xiao, J.; Tian, H.Y.; Wang, M.; Hu, Y.X. Lethal influenza A virus preferentially activates TLR3 and triggers a severe inflammatory response. Virus Res. 2018, 257, 102–112. [Google Scholar] [CrossRef]
- Le Goffic, R.; Pothlichet, J.; Vitour, D.; Fujita, T.; Meurs, E.; Chignard, M.; Si-Tahar, M. Cutting Edge: Influenza A virus activates TLR3-dependent inflammatory and RIG-I-dependent antiviral responses in human lung epithelial cells. J. Immunol. 2007, 178, 3368–3372. [Google Scholar] [CrossRef] [Green Version]
- Lund, J.M.; Alexopoulou, L.; Sato, A.; Karow, M.; Adams, N.C.; Gale, N.W.; Iwasaki, A.; Flavell, R.A. Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2004, 101, 5598–5603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Bowen, G.N.; Padden, C.; Cerny, A.; Finberg, R.W.; Newburger, P.E.; Kurt-Jones, E.A. Toll-like receptor-mediated activation of neutrophils by influenza A virus. Blood 2008, 112, 2028–2034. [Google Scholar] [CrossRef]
- De Marcken, M.; Dhaliwal, K.; Danielsen, A.C.; Gautron, A.S.; Dominguez-Villar, M. TLR7 and TLR8 activate distinct pathways in monocytes during RNA virus infection. Sci. Signal 2019, 12, eaaw1347. [Google Scholar] [CrossRef]
- Ablasser, A.; Poeck, H.; Anz, D.; Berger, M.; Schlee, M.; Kim, S.; Bourquin, C.; Goutagny, N.; Jiang, Z.; Fitzgerald, K.A.; et al. Selection of molecular structure and delivery of RNA oligonucleotides to activate TLR7 versus TLR8 and to induce high amounts of IL-12p70 in primary human monocytes. J. Immunol. 2009, 182, 6824–6833. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Sun, Y.J.; Chen, F.H. Antiviral role of toll-like receptors and cytokines against the new 2009 H1N1 virus infection. Mol. Biol. Rep. 2012, 39, 1163–1172. [Google Scholar] [CrossRef]
- Ablasser, A.; Hur, S. Regulation of cGAS- and RLR-mediated immunity to nucleic acids. Nat. Immunol. 2020, 21, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Rehwinkel, J.; Gack, M.U. RIG-I-like receptors: Their regulation and roles in RNA sensing. Nat. Rev. Immunol. 2020, 20, 537–551. [Google Scholar] [CrossRef]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Naslund, T.I.; Liljestrom, P.; Weber, F.; Sousa, C.R.E. RIG-I-mediated antiviral responses to single-stranded RNA bearing 5’-phosphates. Science 2006, 314, 997–1001. [Google Scholar] [CrossRef] [Green Version]
- Hrincius, E.R.; Dierkes, R.; Anhlan, D.; Wixler, V.; Ludwig, S.; Ehrhardt, C. Phosphatidylinositol-3-kinase (PI3K) is activated by influenza virus vRNA via the pathogen pattern receptor Rig-I to promote efficient type I interferon production. Cell. Microbiol. 2011, 13, 1907–1919. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Q.; Lu, Y.; Raman, S.N.T.; Xu, F.; Wu, Q.; Li, Z.B.; Brownlie, R.; Liu, Q.; Zhou, Y. Nuclear-resident RIG-I senses viral replication inducing antiviral immunity. Nat. Commun. 2018, 9, 3199. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, L.; Barchet, W.; Gilfillan, S.; Cella, M.; Beutler, B.; Flavell, R.A.; Diamond, M.S.; Colonna, M. Essential role of mda-5 in type I IFN responses to polyriboinosinic:polyribocytidylic acid and encephalomyocarditis picornavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 8459–8464. [Google Scholar] [CrossRef] [Green Version]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Rehwinkel, J.; Kato, H.; Takeuchi, O.; Akira, S.; Way, M.; Schiavo, G.; Sousa, C.R.E. Activation of MDA5 Requires Higher-Order RNA Structures Generated during Virus Infection. J. Virol. 2009, 83, 10761–10769. [Google Scholar] [CrossRef] [Green Version]
- Benitez, A.A.; Panis, M.; Xue, J.; Varble, A.; Shim, J.V.; Frick, A.L.; Lopez, C.B.; Sachs, D.; tenOever, B.R. In Vivo RNAi Screening Identifies MDA5 as a Significant Contributor to the Cellular Defense against Influenza A Virus. Cell. Rep. 2015, 11, 1714–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, H.; Takeuchi, O.; Mikamo-Satoh, E.; Hirai, R.; Kawai, T.; Matsushita, K.; Hiiragi, A.; Dermody, T.S.; Fujita, T.; Akira, S. Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5. J. Exp. Med. 2008, 205, 1601–1610. [Google Scholar] [CrossRef]
- Zust, R.; Cervantes-Barragan, L.; Habjan, M.; Maier, R.; Neuman, B.W.; Ziebuhr, J.; Szretter, K.J.; Baker, S.C.; Barchet, W.; Diamond, M.S.; et al. Ribose 2’-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5. Nat. Immunol. 2011, 12, U137–U146. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Wei, F.; Zhang, Y.; Wang, T.; Gao, W.; Yu, S.; Sun, H.; Pu, J.; Sun, Y.; Wang, M.; et al. IFI16 directly senses viral RNA and enhances RIG-I transcription and activation to restrict influenza virus infection. Nat. Microbiol. 2021, 6, 932–945. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Cho, S.W.; Park, P. The Superiority of Ifn-Lambda as a Therapeutic Candidate to Control Acute Influenza Viral Lung Infection. Cytokine 2016, 87, 79–80. [Google Scholar]
- Kim, H.J.; Kim, C.H.; Ryu, J.H.; Kim, M.J.; Park, C.Y.; Lee, J.M.; Holtzman, M.J.; Yoon, J.H. Reactive Oxygen Species Induce Antiviral Innate Immune Response through IFN-lambda Regulation in Human Nasal Epithelial Cells. Am. J. Resp. Cell. Mol. 2013, 49, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.R.; Oberley-Deegan, R.; Wang, S.L.; Nikrad, M.; Funk, C.J.; Hartshorn, K.L.; Mason, R.J. Differentiated Human Alveolar Type II Cells Secrete Antiviral IL-29 (IFN-lambda 1) in Response to Influenza A Infection. J. Immunol. 2009, 182, 1296–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterlund, P.I.; Pietilae, T.E.; Veckman, V.; Kotenko, S.V.; Julkunen, I. IFN regulatory factor family members differentially regulate the expression of type III IFN (IFN-lambda) genes. J. Immunol. 2007, 179, 3434–3442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, M.B.; Ank, N.; Melchjorsen, J.; Paludan, S.R. Expression of Type III Interferon (IFN) in the Vaginal Mucosa Is Mediated Primarily by Dendritic Cells and Displays Stronger Dependence on NF-kappa B than Type I IFNs. J. Virol. 2010, 84, 4579–4586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, M.B.; Paludan, S.R. Mechanisms of Type III Interferon Expression. J. Interf. Cytok Res. 2010, 30, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Thomson, S.J.; Goh, F.G.; Banks, H.; Krausgruber, T.; Kotenko, S.V.; Foxwell, B.M.; Udalova, I.A. The role of transposable elements in the regulation of IFN-lambda1 gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11564–11569. [Google Scholar] [CrossRef] [Green Version]
- Ramos, I.; Smith, G.; Ruf-Zamojski, F.; Martinez-Romero, C.; Fribourg, M.; Carbajal, E.A.; Hartmann, B.M.; Nair, V.D.; Marjanovic, N.; Monteagudo, P.L.; et al. Innate Immune Response to Influenza Virus at Single-Cell Resolution in Human Epithelial Cells Revealed Paracrine Induction of Interferon Lambda 1. J. Virol. 2019, 93, e00559-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazear, H.M.; Nice, T.J.; Diamond, M.S. Interferon-lambda: Immune Functions at Barrier Surfaces and Beyond. Immunity 2015, 43, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Shen, M.L.; Shah, N.K.; Langer, J.A.; Sheikh, F.; Dickensheets, H.; Donnelly, R.P. IFN-lambda s mediate antiviral protection through a distinct class II cytokine receptor complex. Nat. Immunol. 2003, 4, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Kanneganti, T.D.; Franchi, L.; Nunez, G. Caspase-1 inflammasomes in infection and inflammation. J. Leukoc. Biol. 2007, 82, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Sollberger, G.; Strittmatter, G.E.; Kistowska, M.; French, L.E.; Beer, H.D. Caspase-4 Is Required for Activation of Inflammasomes. J. Immunol. 2012, 188, 1992–2000. [Google Scholar] [CrossRef]
- Broz, P.; Monack, D.M. Noncanonical inflammasomes: Caspase-11 activation and effector mechanisms. PLoS Pathog. 2013, 9, e1003144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.G.; Dash, P.; Aldridge, J.R., Jr.; Ellebedy, A.H.; Reynolds, C.; Funk, A.J.; Martin, W.J.; Lamkanfi, M.; Webby, R.J.; Boyd, K.L.; et al. The intracellular sensor NLRP3 mediates key innate and healing responses to influenza A virus via the regulation of caspase-1. Immunity 2009, 30, 566–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, T.D.; Body-Malapel, M.; Amer, A.; Park, J.H.; Whitfield, J.; Franchi, L.; Taraporewala, Z.F.; Miller, D.; Patton, J.T.; Inohara, N.; et al. Critical role for Cryopyrin/Nalp3 in activation of caspase-1 in response to viral infection and double-stranded RNA. J. Biol. Chem. 2006, 281, 36560–36568. [Google Scholar] [CrossRef] [Green Version]
- Kesavardhana, S.; Samir, P.; Zheng, M.; Malireddi, R.K.S.; Karki, R.; Sharma, B.R.; Place, D.E.; Briard, B.; Vogel, P.; Kanneganti, T.D. DDX3X coordinates host defense against influenza virus by activating the NLRP3 inflammasome and type I interferon response. J. Biol. Chem. 2021, 296, 100579. [Google Scholar] [CrossRef]
- Orning, P.; Weng, D.; Starheim, K.; Ratner, D.; Best, Z.; Lee, B.; Brooks, A.; Xia, S.Y.; Wu, H.; Kelliher, M.A.; et al. Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death. Science 2018, 362, 1064–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarhan, J.; Liu, B.C.; Muendlein, H.I.; Li, P.; Nilson, R.; Tang, A.Y.; Rongvaux, A.; Bunnell, S.C.; Shao, F.; Green, D.R.; et al. Caspase-8 induces cleavage of gasdermin D to elicit pyroptosis during Yersinia infection. Proc. Natl. Acad. Sci. USA 2018, 115, E10888–E10897. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Gao, W.Q.; Shi, X.Y.; Ding, J.J.; Liu, W.; He, H.B.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Karki, R.; Vogel, P.; Kanneganti, T.D. Caspase-6 Is a Key Regulator of Innate Immunity, Inflammasome Activation, and Host Defense. Cell 2020, 181, 674–687. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, T.; Man, S.M.; Malireddi, R.K.S.; Karki, R.; Kesavardhana, S.; Place, D.E.; Neale, G.; Vogel, P.; Kanneganti, T.D. ZBP1/DAI is an innate sensor of influenza virus triggering the NLRP3 inflammasome and programmed cell death pathways. Sci. Immunol. 2016, 1, aag2045. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Yin, C.R.; Boyd, D.F.; Quarato, G.; Ingram, J.P.; Shubina, M.; Ragan, K.B.; Ishizuka, T.; Crawford, J.C.; Tummers, B.; et al. Influenza Virus Z-RNAs Induce ZBP1-Mediated Necroptosis. Cell 2020, 180, 1115–1129. [Google Scholar] [CrossRef]
- Spackman, E. A Brief Introduction to Avian Influenza Virus. Methods Mol. Biol. 2014, 1161, 61–68. [Google Scholar]
- Jiao, P.R.; Song, H.; Liu, X.K.; Song, Y.F.; Cui, J.; Wu, S.Y.; Ye, J.Q.; Qu, N.A.; Zhang, T.M.; Liao, M. Pathogenicity, Transmission and Antigenic Variation of H5N1 Highly Pathogenic Avian Influenza Viruses. Front. Microbiol. 2016, 7, 635. [Google Scholar] [CrossRef] [Green Version]
- Downing, T.; Lloyd, A.T.; O’Farrelly, C.; Bradley, D.G. The Differential Evolutionary Dynamics of Avian Cytokine and TLR Gene Classes. J. Immunol. 2010, 184, 6993–7000. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Cheng, A.C.; Wang, M.S. Innate sensing of viruses by pattern recognition receptors in birds. Vet. Res. 2013, 44, 82. [Google Scholar] [CrossRef] [Green Version]
- Cormican, P.; Lloyd, A.T.; Downing, T.; Connell, S.J.; Bradley, D.; O’Farrelly, C. The avian Toll-Like receptor pathway—Subtle differences amidst general conformity. Dev. Comp. Immunol. 2009, 33, 967–973. [Google Scholar] [CrossRef]
- Karpala, A.J.; Lowenthal, J.W.; Bean, A.G. Activation of the TLR3 pathway regulates IFNbeta production in chickens. Dev. Comp. Immunol. 2008, 32, 435–444. [Google Scholar] [CrossRef]
- Ahmed-Hassan, H.; Abdul-Cader, M.S.; Sabry, M.A.; Hamza, E.; Sharif, S.; Nagy, E.; Abdul-Careem, M.F. Double-Stranded Ribonucleic Acid-Mediated Antiviral Response against Low Pathogenic Avian Influenza Virus Infection. Viral Immunol. 2018, 31, 433–446. [Google Scholar] [CrossRef] [PubMed]
- St Paul, M.; Mallick, A.I.; Read, L.R.; Villanueva, A.I.; Parvizi, P.; Abdul-Careem, M.F.; Nagy, E.; Sharif, S. Prophylactic treatment with Toll-like receptor ligands enhances host immunity to avian influenza virus in chickens. Vaccine 2012, 30, 4524–4531. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, M.; Shamovsky, V.; D’Eustachio, P. Human and chicken TLR pathways: Manual curation and computer-based orthology analysis. Mamm. Genome 2011, 22, 130–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philbin, V.J.; Iqbal, M.; Boyd, Y.; Goodchild, M.J.; Beal, R.K.; Bumstead, N.; Young, J.; Smith, A.L. Identification and characterization of a functional, alternatively spliced Toll-like receptor 7 (TLR7) and genomic disruption of TLR8 in chickens. Immunology 2005, 114, 507–521. [Google Scholar] [CrossRef]
- Brownlie, R.; Zhu, J.Z.; Allan, B.; Mutwiri, G.K.; Babiuk, L.A.; Potter, A.; Griebel, P. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides. Mol. Immunol. 2009, 46, 3163–3170. [Google Scholar] [CrossRef]
- Abdul-Cader, M.S.; De Silva Senapathi, U.; Nagy, E.; Sharif, S.; Abdul-Careem, M.F. Antiviral response elicited against avian influenza virus infection following activation of toll-like receptor (TLR)7 signaling pathway is attributable to interleukin (IL)-1beta production. BMC Res. Notes 2018, 11, 859. [Google Scholar] [CrossRef]
- Campbell, L.K.; Magor, K.E. Pattern Recognition Receptor Signaling and Innate Responses to Influenza A Viruses in the Mallard Duck, Compared to Humans and Chickens. Front. Cell Infect. Microbiol. 2020, 10, 209. [Google Scholar] [CrossRef]
- Barber, M.R.W.; Aldridge, J.R.; Fleming-Canepa, X.; Wang, Y.D.; Webster, R.G.; Magor, K.E. Identification of avian RIG-I responsive genes during influenza infection. Mol. Immunol. 2013, 54, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Karpala, A.J.; Stewart, C.; McKay, J.; Lowenthal, J.W.; Bean, A.G. Characterization of chicken Mda5 activity: Regulation of IFN-beta in the absence of RIG-I functionality. J. Immunol. 2011, 186, 5397–5405. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yu, D.D.; Fan, Y.; Liu, Y.P.; Yao, Y.G. Evolutionary selection on MDA5 and LGP2 in the chicken preserves antiviral competence in the absence of RIG-I. J. Genet. Genom. 2019, 46, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.Q.; Sun, Y.J.; Wang, H.A.; Yan, Y.X.; Ding, C.; Sun, J.H. Chicken STING Mediates Activation of the IFN Gene Independently of the RIG-I Gene. J. Immunol. 2015, 195, 3922–3936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liniger, M.; Summerfield, A.; Zimmer, G.; McCullough, K.C.; Ruggli, N. Chicken Cells Sense Influenza A Virus Infection through MDA5 and CARDIF Signaling Involving LGP2. J. Virol. 2012, 86, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.L.; Li, Y.F.; Qi, W.B.; Ma, X.L.; Yu, K.X.; Huang, B.; Liao, M.; Li, F.; Pan, J.; Song, M.X. Comparative analysis of selected innate immune-related genes following infection of immortal DF-1 cells with highly pathogenic (H5N1) and low pathogenic (H9N2) avian influenza viruses. Virus Genes. 2015, 50, 189–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.X.; Cheng, Y.Q.; Shan, W.Y.; Ma, J.J.; Wang, H.A.; Sun, J.H.; Yan, Y.X. Chicken interferon regulatory factor 1 (IRF1) involved in antiviral innate immunity via regulating IFN-beta production. Dev. Comp. Immunol. 2018, 88, 77–82. [Google Scholar] [CrossRef]
- Cheng, Y.Q.; Zhu, W.X.; Ding, C.; Niu, Q.N.; Wang, H.G.; Yan, Y.X.; Sun, J.H. IRF7 Is Involved in Both STING and MAVS Mediating IFN- beta Signaling in IRF3-Lacking Chickens. J. Immunol. 2019, 203, 1930–1942. [Google Scholar] [CrossRef]
- Niu, Q.; Cheng, Y.; Wang, H.; Yan, Y.; Sun, J. Chicken DDX3X Activates IFN-beta via the chSTING-chIRF7-IFN-beta Signaling Axis. Front. Immunol. 2019, 10, 822. [Google Scholar] [CrossRef]
- Huo, H.; Wang, Y.; Wang, D.F.; Wang, Y.P.; Chen, X.H.; Zhao, L.L.; Chen, H.Y. Duck RIG-I restricts duck enteritis virus infection. Vet. Microbiol. 2019, 230, 78–85. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, Z.Y.; Wang, B.; Yu, Q.M.; Liu, R.; Xu, Q.; Chang, G.B.; Ding, J.T.; Chen, G.H. Duck RIG-I CARD Domain Induces the Chicken IFN-beta by Activating NF-kappa B. Biomed. Res. Int. 2015, 2015, 348792. [Google Scholar]
- Shao, Q.; Xu, W.P.; Yan, L.; Liu, J.H.; Rui, L.; Xiao, X.; Yu, X.X.; Lu, Y.N.; Li, Z.D. Function of duck RIG-I in induction of antiviral response against IBDV and avian influenza virus on chicken cells. Virus Res. 2014, 191, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.Q.; Huang, Q.Q.; Ji, W.H.; Du, B.; Fu, Q.; An, H.T.; Li, J.; Wang, H.G.; Yan, Y.X.; Ding, C.; et al. Muscovy duck retinoic acid-induced gene I (MdRIG-I) functions in innate immunity against H9N2 avian influenza viruses (AIV) infections. Vet. Immunol. Immunopathol. 2015, 163, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Cui, J.; Song, Y.; Zhang, S.; Han, F.; Yuan, R.; Gong, L.; Jiao, P.; Liao, M. Duck MDA5 functions in innate immunity against H5N1 highly pathogenic avian influenza virus infections. Vet. Res. 2014, 45, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.N.; Wu, S.Y.; Wu, W.B.; Liang, Y.W.; Zhuang, H.B.; Ye, Z.Y.; Qu, X.Y.; Liao, M.; Jiao, P.R. The Biological Characteristics of Novel H5N6 Highly Pathogenic Avian Influenza Virus and Its Pathogenesis in Ducks. Front. Microbiol. 2021, 12, 628545. [Google Scholar] [CrossRef]
- Anderson, T.K.; Chang, J.; Arendsee, Z.W.; Venkatesh, D.; Souza, C.K.; Kimble, J.B.; Lewis, N.S.; Davis, C.T.; Vincent, A.L. Swine Influenza A Viruses and the Tangled Relationship with Humans. Cold Spring Harb. Perspect. Med. 2021, 11, a038737. [Google Scholar] [CrossRef] [Green Version]
- Epidemiology, G.D.; Org, P.A.H.; Org, W.H.; Centers for Disease Control and Prevention (CDC). Outbreak of Swine-Origin Influenza A (H1N1) Virus Infection-Mexico, March-April 2009. MMWR Morb. Mortal. Wkly. Rep. 2009, 301, 2321–2322. [Google Scholar]
- Zhang, J.Q.; Miao, J.F.; Hou, J.B.; Lu, C.P. The effects of H3N2 swine influenza virus infection on TLRs and RLRs signaling pathways in porcine alveolar macrophages. Virol. J. 2015, 12, 61. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Ortega, M.; Melo, S.; Punyadarsaniya, D.; Rame, C.; Olivier, M.; Soubieux, D.; Marc, D.; Simon, G.; Herrler, G.; Berri, M.; et al. Innate immune response to a H3N2 subtype swine influenza virus in newborn porcine trachea cells, alveolar macrophages, and precision-cut lung slices. Vet. Res. 2014, 45, 42. [Google Scholar] [CrossRef] [Green Version]
- Husser, L.; Alves, M.P.; Ruggli, N.; Summerfield, A. Identification of the role of RIG-I, MDA-5 and TLR3 in sensing RNA viruses in porcine epithelial cells using lentivirus-driven RNA interference. Virus Res. 2011, 159, 9–16. [Google Scholar] [CrossRef]
- Haque, M.M.; Murale, D.P.; Lee, J.S. Role of microRNA and Oxidative Stress in Influenza A Virus Pathogenesis. Int. J. Mol. Sci. 2020, 21, 8962. [Google Scholar] [CrossRef]
- Guo, M.; Li, F.B.; Ji, J.S.; Liu, Y.F.; Liu, F.; Zhao, Y.Y.; Li, J.H.; Han, S.; Wang, Q.X.; Ding, G.S. Inhibition of miR-93 promotes interferon effector signaling to suppress influenza A infection by upregulating JAK1. Int. Immunopharmacol. 2020, 86, 106754. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.Z.; Zhang, X.H.; Wu, Z.; Huang, K.; Sun, X.M.; Chen, H.C.; Jin, M.L. The Downregulation of MicroRNA hsa-miR-340-5p in IAV-Infected A549 Cells Suppresses Viral Replication by Targeting RIG-I and OAS2. Mol. Ther. Nucleic Acids 2019, 14, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.Q.; Yan, Y.; Sen Tan, K.; Liu, J.; Chow, V.T.; Tao, Z.Z.; Wang, D.Y. MicroRNA-146a induction during influenza H3N2 virus infection targets and regulates TRAF6 levels in human nasal epithelial cells (hNECs). Exp. Cell Res. 2017, 352, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dong, C.; Sun, X.; Li, Z.; Zhang, M.; Guan, Z.; Duan, M. Induction of the cellular miR-29c by influenza virus inhibits the innate immune response through protection of A20 mRNA. Biochem. Biophys. Res. Commun. 2014, 450, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.M.; Ouyang, J.; Wei, J.Y.; Maarouf, M.; Chen, J.L. Involvement of Host Non-Coding RNAs in the Pathogenesis of the Influenza Virus. Int. J. Mol. Sci. 2017, 18, 39. [Google Scholar] [CrossRef] [Green Version]
- Jureka, A.S.; Kleinpeter, A.B.; Tipper, J.L.; Harrod, K.S.; Petit, C.M. The influenza NS1 protein modulates RIG-I activation via a strain-specific direct interaction with the second CARD of RIG-I. J. Biological Chem. 2020, 295, 1153–1164. [Google Scholar] [CrossRef]
- Gack, M.U.; Albrecht, R.A.; Urano, T.; Inn, K.S.; Huang, I.C.; Carnero, E.; Farzan, M.; Inoue, S.; Jung, J.U.; Garcia-Sastre, A. Influenza A Virus NS1 Targets the Ubiquitin Ligase TRIM25 to Evade Recognition by the Host Viral RNA Sensor RIG-I. Cell Host Microbe 2009, 5, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Rajsbaum, R.; Albrecht, R.A.; Wang, M.K.; Maharaj, N.P.; Versteeg, G.A.; Nistal-Villan, E.; Garcia-Sastre, A.; Gack, M.U. Species-Specific Inhibition of RIG-I Ubiquitination and IFN Induction by the Influenza A Virus NS1 Protein. PLoS Pathog. 2012, 8, e1003059. [Google Scholar] [CrossRef]
- Koliopoulos, M.G.; Lethier, M.; van der Veen, A.G.; Haubrich, K.; Hennig, J.; Kowalinski, E.; Stevens, R.V.; Martin, S.R.; Sousa, C.R.E.; Cusack, S.; et al. Molecular mechanism of influenza A NS1-mediated TRIM25 recognition and inhibition. Nat. Commun. 2018, 9, 1820. [Google Scholar] [CrossRef]
- Moriyama, M.; Chen, I.Y.; Kawaguchi, A.; Koshiba, T.; Nagata, K.; Takeyama, H.; Hasegawa, H.; Ichinohe, T. The RNA- and TRIM25-Binding Domains of Influenza Virus NS1 Protein Are Essential for Suppression of NLRP3 Inflammasome-Mediated Interleukin-1 beta Secretion. J. Virol. 2016, 90, 4105–4114. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Peng, H.; Jiang, W.; Song, L. NS1 protein of avian influenza A virus prevents activation of NF-kappa B through binding to IKK alpha and IKK beta. Int. J. Infect. Dis. 2010, 14, E82–E83. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.J.; Song, L.P.; Li, J.D.; Zhang, Z.Z.; Peng, H.Y.; Jiang, W.; Wang, Q.T.; Kang, T.B.; Chen, S.; Huang, W.L. Influenza A virus-encoded NS1 virulence factor protein inhibits innate immune response by targeting IKK. Cell. Microbiol. 2012, 14, 1849–1866. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Shih, M.C.; Chang, H.C.; Lin, K.J.; Chen, L.F.; Huang, S.W.; Yang, M.L.; Ma, S.K.; Shiau, A.L.; Wang, J.R.; et al. Influenza a virus NS1 resembles a TRAF3-interacting motif to target the RNA sensing-TRAF3-type I IFN axis and impair antiviral innate immunity. J. Biomed. Sci. 2021, 28, 66. [Google Scholar] [CrossRef]
- Wolf, J.J.; Xia, C.; Studstill, C.J.; Ngo, H.; Brody, S.L.; Anderson, P.E.; Hahm, B. Influenza A virus NS1 induces degradation of sphingosine 1-phosphate lyase to obstruct the host innate immune response. Virology 2021, 558, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Yoshizumi, T.; Ichinohe, T.; Sasaki, O.; Otera, H.; Kawabata, S.I.; Mihara, K.; Koshiba, T. Influenza A virus protein PB1-F2 translocates into mitochondria via Tom40 channels and impairs innate immunity. Nat. Commun. 2014, 5, 4713. [Google Scholar] [CrossRef]
- Varga, Z.T.; Grant, A.; Manicassamy, B.; Palese, P. Influenza Virus Protein PB1-F2 Inhibits the Induction of Type I Interferon by Binding to MAVS and Decreasing Mitochondrial Membrane Potential. J. Virol. 2012, 86, 8359–8366. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.F.; Zhu, Y.X.; Ren, C.W.; Yang, S.K.; Tian, S.; Chen, H.C.; Jin, M.L.; Zhou, H.B. Influenza A virus protein PB1-F2 impairs innate immunity by inducing mitophagy. Autophagy 2021, 17, 496–511. [Google Scholar] [CrossRef]
- Allison, H.; Holdsworth, G.; McNamara, L.M. Scl-Ab reverts pro-osteoclastogenic signalling and resorption in estrogen deficient osteocytes. BMC Mol. Cell Biol. 2020, 21, 78. [Google Scholar] [CrossRef]
- Dudek, S.E.; Wixler, L.; Nordhoff, C.; Nordmann, A.; Anhlan, D.; Wixler, V.; Ludwig, S. The influenza virus PB1-F2 protein has interferon antagonistic activity. Biol. Chem. 2011, 392, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Xu, S.; Wei, Y.L.; Zhang, X.G.; Wang, Q.; Jia, Y.N.; Wang, W.B.; Han, L.; Chen, Z.S.; Wang, Z.X.; et al. The PB1 protein of influenza A virus inhibits the innate immune response by targeting MAVS for NBR1-mediated selective autophagic degradation. PLoS Pathog. 2021, 17, e1009300. [Google Scholar] [CrossRef]
- Hayashi, T.; MacDonald, L.A.; Takimoto, T. Influenza A Virus Protein PA-X Contributes to Viral Growth and Suppression of the Host Antiviral and Immune Responses. J. Virol. 2015, 89, 6442–6452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigby, R.E.; Wise, H.M.; Smith, N.; Digard, P.; Rehwinkel, J. PA-X antagonises MAVS-dependent accumulation of early type I interferon messenger RNAs during influenza A virus infection. Sci. Rep. 2019, 9, 7216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Qi, W.; Chang, Q.; Chen, R.; Zhen, D.; Liao, M.; Wen, J.; Deng, Y. Influenza A virus protein PA-X suppresses host Ankrd17-mediated immune responses. Microbiol. Immunol. 2021, 65, 48–59. [Google Scholar] [CrossRef]
- Hu, J.; Kong, M.; Cui, Z.; Gao, Z.; Ma, C.X.; Hu, Z.L.; Jiao, X.N.; Liu, X.F. PA-X protein of H5N1 avian influenza virus inhibits NF-kappaB activity, a potential mechanism for PA-X counteracting the host innate immune responses. Vet. Microbiol. 2020, 250, 108838. [Google Scholar] [CrossRef] [PubMed]
- Liniger, M.; Moulin, H.R.; Sakoda, Y.; Ruggli, N.; Summerfield, A. Highly pathogenic avian influenza virus H5N1 controls type I IFN induction in chicken macrophage HD-11 cells: A polygenic trait that involves NS1 and the polymerase complex. Virol. J. 2012, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, J.; Smith, N.; Ross, C.; Iqbal, M.; Goodbourn, S.; Digard, P.; Barclay, W.S.; Shelton, H. The cellular localization of avian influenza virus PB1-F2 protein alters the magnitude of IFN2 promoter and NFkappaB-dependent promoter antagonism in chicken cells. J. Gen Virol. 2019, 100, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Evseev, D.; Stevens, C.A.; Moghrabi, A.; Miranzo-Navarro, D.; Fleming-Canepa, X.; Tetrault, D.G.; Magor, K.E. Influenza PB1-F2 Inhibits Avian MAVS Signaling. Viruses 2020, 12, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.S.; Liu, G.; Thulasi Raman, S.N.; Landreth, S.L.; Liu, Q.; Zhou, Y. NS1 Protein of 2009 Pandemic Influenza A Virus Inhibits Porcine NLRP3 Inflammasome-Mediated Interleukin-1 Beta Production by Suppressing ASC Ubiquitination. J. Virol. 2018, 92, e00022-18. [Google Scholar] [CrossRef] [Green Version]
- Cleri, D.J.; Ricketti, A.J.; Vernaleo, J.R. Severe Acute Respiratory Syndrome (SARS). Infect. Dis. Clin. N. Am. 2010, 24, 175–202. [Google Scholar] [CrossRef]
- Mousavizadeh, L.; Ghasemi, S. Review Article Genotype and phenotype of COVID-19: Their roles in pathogenesis. J. Microbiol. Immunol. 2021, 54, 159–163. [Google Scholar]
- Chen, Y.; Liu, Q.Y.; Guo, D.Y. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.D.; Sun, Y.H.; Zhou, J.M.; Ye, Q. The Global Epidemic of the SARS-CoV-2 Delta Variant, Key Spike Mutations and Immune Escape. Front. Immunol. 2021, 12, 751778. [Google Scholar] [CrossRef]
- Karim, S.S.A.; Karim, Q.A. Omicron SARS-CoV-2 variant: A new chapter in the COVID-19 pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef]
- Spiegel, M.; Pichlmair, A.; Martinez-Sobrido, L.; Cros, J.; Garcia-Sastre, A.; Haller, O.; Weber, F. Inhibition of beta interferon induction by severe acute respiratory syndrome coronavirus suggests a two-step model for activation of interferon regulatory factor 3. J. Virol. 2005, 79, 2079–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wathelet, M.G.; Orr, M.; Frieman, M.B.; Baric, R.S. Severe acute respiratory syndrome coronavirus evades antiviral signaling: Role of nsp1 and rational design of an attenuated strain. J. Virol. 2007, 81, 11620–11633. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045. [Google Scholar] [CrossRef]
- Versteeg, G.A.; Bredenbeek, P.J.; van den Worm, S.H.E.; Spaan, W.J.M. Group 2 coronaviruses prevent immediate early interferon induction by protection of viral RNA from host cell recognition. Virology 2007, 361, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Channappanavar, R.; Kanneganti, T.D. Coronaviruses: Innate Immunity, Inflammasome Activation, Inflammatory Cell Death, and Cytokines. Trends Immunol. 2020, 41, 1083–1099. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Gargan, S.; Lu, Y.X.; Stevenson, N.J. An Overview of Current Knowledge of Deadly CoVs and Their Interface with Innate Immunity. Viruses 2021, 13, 560. [Google Scholar] [CrossRef] [PubMed]
- Totura, A.L.; Whitmore, A.; Agnihothram, S.; Schafer, A.; Katze, M.G.; Heise, M.T.; Baric, R.S. Toll-Like Receptor 3 Signaling via TRIF Contributes to a Protective Innate Immune Response to Severe Acute Respiratory Syndrome Coronavirus Infection. mBio 2015, 6, e00638-15. [Google Scholar] [CrossRef] [Green Version]
- Bortolotti, D.; Gentili, V.; Rizzo, S.; Schiuma, G.; Beltrami, S.; Strazzabosco, G.; Fernandez, M.; Caccuri, F.; Caruso, A.; Rizzo, R. TLR3 and TLR7 RNA Sensor Activation during SARS-CoV-2 Infection. Microorganisms 2021, 9, 1820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bastard, P.; Liu, Z.Y.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef]
- Croci, S.; Venneri, M.A.; Mantovani, S.; Fallerini, C.; Benetti, E.; Picchiotti, N.; Campolo, F.; Imperatore, F.; Palmieri, M.; Daga, S.; et al. The polymorphism L412F in TLR3 inhibits autophagy and is a marker of severe COVID-19 in males. Autophagy 2021, 18, 1662–1672. [Google Scholar] [CrossRef]
- Dyavar, S.R.; Singh, R.; Emani, R.; Pawar, G.P.; Chaudhari, V.D.; Podany, A.T.; Avedissian, S.N.; Fletcher, C.V.; Salunke, D.B. Role of toll-like receptor 7/8 pathways in regulation of interferon response and inflammatory mediators during SARS-CoV2 infection and potential therapeutic options. Biomed. Pharmacother. 2021, 141, 111794. [Google Scholar] [CrossRef]
- Khanmohammadi, S.; Rezaei, N. Role of Toll-like receptors in the pathogenesis of COVID-19. J. Med. Virol. 2021, 93, 2735–2739. [Google Scholar] [CrossRef]
- Cervantes-Barragan, L.; Zust, R.; Weber, F.; Spiegel, M.; Lang, K.S.; Akira, S.; Thiel, V.; Ludewig, B. Control of coronavirus infection through plasmacytoid dendritic-cell-derived type I interferon. Blood 2007, 109, 1131–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Channappanavar, R.; Fehr, A.R.; Zheng, J.; Wohlford-Lenane, C.; Abrahante, J.E.; Mack, M.; Sompallae, R.; McCray, P.B., Jr.; Meyerholz, D.K.; Perlman, S. IFN-I response timing relative to virus replication determines MERS coronavirus infection outcomes. J. Clin. Investig. 2019, 129, 3625–3639. [Google Scholar] [CrossRef]
- Moreno-Eutimio, M.A.; Lopez-Macias, C.; Pastelin-Palacios, R. Bioinformatic analysis and identification of single-stranded RNA sequences recognized by TLR7/8 in the SARS-CoV-2, SARS-CoV, and MERS-CoV genomes. Microbes Infect. 2020, 22, 226–229. [Google Scholar] [CrossRef]
- Fallerini, C.; Daga, S.; Mantovani, S.; Benetti, E.; Picchiotti, N.; Francisci, D.; Paciosi, F.; Schiaroli, E.; Baldassarri, M.; Fava, F.; et al. Association of Toll-like receptor 7 variants with life-threatening COVID-19 disease in males: Findings from a nested case-control study. eLife 2021, 10, e67569. [Google Scholar] [CrossRef] [PubMed]
- Mahallawi, W.H.; Suliman, B.A. TLR8 is highly conserved among the Saudi population and its mutations have no effect on the severity of COVID-19 symptoms. Am. J. Clin. Exp. Immunol. 2021, 10, 71–76. [Google Scholar] [PubMed]
- Conti, P.; Younes, A. Coronavirus COV-19/SARS-CoV-2 affects women less than men: Clinical response to viral infection. J. Biol. Regul. Homeost. Agents 2020, 34, 339–343. [Google Scholar] [PubMed]
- De Groot, N.G.; Bontrop, R.E. COVID-19 pandemic: Is a gender-defined dosage effect responsible for the high mortality rate among males? Immunogenetics 2020, 72, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Riva, L.; Pu, Y.; Martin-Sancho, L.; Kanamune, J.; Yamamoto, Y.; Sakai, K.; Gotoh, S.; Miorin, L.; De Jesus, P.D.; et al. MDA5 Governs the Innate Immune Response to SARS-CoV-2 in Lung Epithelial Cells. Cell Rep. 2021, 34, 108628. [Google Scholar] [CrossRef] [PubMed]
- Rebendenne, A.; Valadão, A.L.C.; Tauziet, M.; Maarifi, G.; Bonaventure, B.; McKellar, J.; Planès, R.; Nisole, S.; Arnaud-Arnould, M.; Moncorgé, O.; et al. SARS-CoV-2 triggers an MDA-5-dependent interferon response which is unable to control replication in lung epithelial cells. J. Virol. 2021, 95, e02415-20. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, N.G.; Chauveau, L.; Hertzog, J.; Bridgeman, A.; Fowler, G.; Moonen, J.P.; Dupont, M.; Russell, R.A.; Noerenberg, M.; Rehwinkel, J. The RNA sensor MDA5 detects SARS-CoV-2 infection. Sci. Rep. 2021, 11, 13638. [Google Scholar] [CrossRef]
- Yang, D.M.; Geng, T.T.; Harrison, A.G.; Wang, P.H. Differential roles of RIG-I like receptors in SARS-CoV-2 infection. Mil. Med. Res. 2021, 8, 49. [Google Scholar] [CrossRef]
- Thorne, L.G.; Reuschl, A.K.; Zuliani-Alvarez, L.; Whelan, M.V.X.; Turner, J.; Noursadeghi, M.; Jolly, C.; Towers, G.J. SARS-CoV-2 sensing by RIG-I and MDA5 links epithelial infection to macrophage inflammation. EMBO J. 2021, 40, e107826. [Google Scholar] [CrossRef]
- Yamada, T.; Sato, S.; Sotoyama, Y.; Orba, Y.; Sawa, H.; Yamauchi, H.; Sasaki, M.; Takaoka, A. RIG-I triggers a signaling-abortive anti-SARS-CoV-2 defense in human lung cells. Nat. Immunol. 2021, 22, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Deinhardt-Emmer, S.; Bottcher, S.; Haring, C.; Giebeler, L.; Henke, A.; Zell, R.; Jungwirth, J.; Jordan, P.M.; Werz, O.; Hornung, F.; et al. SARS-CoV-2 Causes Severe Epithelial Inflammation and Barrier Dysfunction. J. Virol. 2021, 95, e00110-21. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.W.; Wang, T.; Zhang, X.Y.; Chen, H.L.; Yu, H.J.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; Su, W.R.; Tang, H.; Le, W.Q.; Zhang, X.P.; Zheng, Y.F.; Liu, X.X.; Xie, L.H.; Li, J.M.; Ye, J.G.; et al. Immune cell profiling of COVID-19 patients in the recovery stage by single-cell sequencing. Cell Discov. 2020, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.Y.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, D.F.; te Velde, A.A. Severe COVID-19: NLRP3 Inflammasome Dysregulated. Front. Immunol. 2020, 11, 1580. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; de Sa, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Goncalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Wu, H.B.; Yao, X.H.; Zhang, D.Y.; Zhou, Y.G.; Fu, B.Q.; Wang, W.; Li, H.; Wang, Z.; Hu, Z.M.; et al. Pyroptotic macrophages stimulate the SARS-CoV-2-associated cytokine storm. Cell. Mol. Immunol. 2021, 18, 1305–1307. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.R.; To, R.K.; Hanna, J.; Spector, S.A. SARS-CoV-2, SARS-CoV-1, and HIV-1 derived ssRNA sequences activate the NLRP3 inflammasome in human macrophages through a non-classical pathway. iScience 2021, 24, 102295. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Williams, E.P.; Malireddi, R.K.S.; Karki, R.; Banoth, B.; Burton, A.; Webby, R.; Channappanavar, R.; Jonsson, C.B.; Kanneganti, T.D. Impaired NLRP3 inflammasome activation/pyroptosis leads to robust inflammatory cell death via caspase-8/RIPK3 during coronavirus infection. J. Biol. Chem. 2020, 295, 14040–14052. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chu, H.; Wong, B.H.; Chiu, M.C.; Wang, D.; Li, C.; Liu, X.; Yang, D.; Poon, V.K.; Cai, J.; et al. Activation of C-Type Lectin Receptor and (RIG)-I-Like Receptors Contributes to Proinflammatory Response in Middle East Respiratory Syndrome Coronavirus-Infected Macrophages. J. Infect. Dis. 2020, 221, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, J.; Teng, Y.; Sun, H.; Tian, G.; He, L.; Li, P.; Chen, Y.; Guo, Y.; Li, J.; et al. Complement Receptor C5aR1 Inhibition Reduces Pyroptosis in hDPP4-Transgenic Mice Infected with MERS-CoV. Viruses 2019, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Rabouw, H.H.; Langereis, M.A.; Knaap, R.C.M.; Dalebout, T.J.; Canton, J.; Sola, I.; Enjuanes, L.; Bredenbeek, P.J.; Kikkert, M.; de Groot, R.J.; et al. Middle East Respiratory Coronavirus Accessory Protein 4a Inhibits PKR-Mediated Antiviral Stress Responses. PLoS Pathog. 2016, 12, e1005982. [Google Scholar] [CrossRef] [PubMed]
- Dosch, S.F.; Mahajan, S.D.; Collins, A.R. SARS coronavirus spike protein-induced innate immune response occurs via activation of the NF-kappaB pathway in human monocyte macrophages in vitro. Virus Res. 2009, 142, 19–27. [Google Scholar] [CrossRef]
- Chen, I.Y.; Moriyama, M.; Chang, M.F.; Ichinohe, T. Severe Acute Respiratory Syndrome Coronavirus Viroporin 3a Activates the NLRP3 Inflammasome. Front. Microbiol. 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, A.; Mukherjee, S. In Silico studies on the comparative characterization of the interactions of SARS-CoV-2 spike glycoprotein with ACE-2 receptor homologs and human TLRs. J. Med. Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Renner, D.M.; Comar, C.E.; Whelan, J.N.; Reyes, H.M.; Cardenas-Diaz, F.L.; Truitt, R.; Tan, L.H.; Dong, B.; Alysandratos, K.D.; et al. SARS-CoV-2 induces double-stranded RNA-mediated innate immune responses in respiratory epithelial-derived cells and cardiomyocytes. Proc. Natl. Acad. Sci. USA 2021, 118, e2022643118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, X.; Lei, X.; Xiao, X.; Jiao, T.; Ma, R.; Dong, X.; Jiang, Q.; Wang, W.; Shi, Y.; et al. Sensing of cytoplasmic chromatin by cGAS activates innate immune response in SARS-CoV-2 infection. Signal Transduct. Target Ther. 2021, 6, 382. [Google Scholar] [CrossRef]
- Ren, H.; Ma, C.; Peng, H.; Zhang, B.; Zhou, L.; Su, Y.; Gao, X.; Huang, H. Micronucleus production, activation of DNA damage response and cGAS-STING signaling in syncytia induced by SARS-CoV-2 infection. Biol. Direct. 2021, 16, 20. [Google Scholar] [CrossRef] [PubMed]
- Domizio, J.D.; Gulen, M.F.; Saidoune, F.; Thacker, V.V.; Yatim, A.; Sharma, K.; Nass, T.; Guenova, E.; Schaller, M.; Conrad, C.; et al. The cGAS-STING pathway drives type I IFN immunopathology in COVID-19. Nature 2022, 603, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Raja, A.; Raj, G.D.; Kumanan, K. Emergence of variant avian infectious bronchitis virus in India. Iran. J. Vet. Res. 2020, 21, 33–39. [Google Scholar] [PubMed]
- Zhang, Y.; Xu, Z.C.; Cao, Y.C. Host Antiviral Responses against Avian Infectious Bronchitis Virus (IBV): Focus on Innate Immunity. Viruses 2021, 13, 1698. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xu, S.; Li, X.; Wang, J.; Jiang, Y.; Hu, W.; Ruan, W. Infectious bronchitis virus inhibits activation of the TLR7 pathway, but not the TLR3 pathway. Arch. Virol. 2020, 165, 2037–2043. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xie, Z.; Dai, J.; Cao, Y.; Hou, J.; Zheng, Y.; Wei, T.; Mo, M.; Wei, P. Responses of the Toll-like receptor and melanoma differentiation-associated protein 5 signaling pathways to avian infectious bronchitis virus infection in chicks. Virol. Sin. 2016, 31, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameka, A.M.; Haddadi, S.; Kim, D.S.; Cork, S.C.; Abdul-Careem, M.F. Induction of innate immune response following infectious bronchitis corona virus infection in the respiratory tract of chickens. Virology 2014, 450–451, 114–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.S.; Yang, X.; Wang, H.N.; Zeng, F.Y.; Zhang, Z.K.; Zhang, A.Y.; Liu, H. The establishment and characteristics of cell-adapted IBV strain H120. Arch. Virol. 2016, 161, 3179–3187. [Google Scholar] [CrossRef]
- Yu, L.P.; Zhang, X.R.; Wu, T.Q.; Su, J.; Wang, Y.Y.; Wang, Y.X.; Ruan, B.Y.; Niu, X.S.; Wu, Y.T. Avian infectious bronchitis virus disrupts the melanoma differentiation associated gene 5 (MDA5) signaling pathway by cleavage of the adaptor protein MAVS. BMC Vet. Res. 2017, 13, 332. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.L.; Cui, P.F.; Ni, R.Q.; Gong, H.L.; Li, H.; Yan, W.J.; Fu, X.; Chen, L.; Lei, C.W.; Wang, H.N.; et al. Chicken-Derived Pattern Recognition Receptor chLGP2 Inhibits the Replication and Proliferation of Infectious Bronchitis Virus. Front. Microbiol. 2022, 12, 810215. [Google Scholar] [CrossRef]
- Cao, L.; Ge, X.; Gao, Y.; Ren, Y.; Ren, X.; Li, G. Porcine epidemic diarrhea virus infection induces NF-kappaB activation through the TLR2, TLR3 and TLR9 pathways in porcine intestinal epithelial cells. J. Gen. Virol. 2015, 96, 1757–1767. [Google Scholar] [CrossRef]
- Ding, Z.; An, K.; Xie, L.L.; Wu, W.; Zhang, R.X.; Wang, D.; Fang, Y.; Chen, H.C.; Xiao, S.B.; Fang, L.R. Transmissible gastroenteritis virus infection induces NF-kappa B activation through RLR-mediated signaling. Virology 2017, 507, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yin, L.; Xue, M.; Chen, J.; Li, L.; Fu, F.; Feng, L.; Liu, P. Coronavirus Porcine Deltacoronavirus Upregulates MHC Class I Expression through RIG-I/IRF1-Mediated NLRC5 Induction. J. Virol. 2022, 96, e0015822. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, S.Q.; Wang, H.F.; Wu, Z.C.; Bao, W.B.; Wu, S.L. Effects of porcine epidemic diarrhea virus infection on Toll-like receptor expression and cytokine levels in porcine intestinal epithelial cells. Pol. J. Vet. Sci. 2020, 23, 119–126. [Google Scholar] [PubMed]
- Wei, G.; Luo, S.; Wu, W.; Hu, J.; Zhou, R. Activation of Interleukin-1β Release and Pyroptosis by Transmissible Gastroenteritis Virus Is Dependent on the NOD-Like Receptor Protein 3 Inflammasome in Porcine Intestinal Epithelial Cell Line. Viral Immunol. 2021, 34, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, M.N.; Aydemir, H.B.; Korkmaz, E.M.; Budak, M.; Cekin, N.; Pinarbasi, E. Computationally predicted SARS-CoV-2 encoded microRNAs target NFKB, JAK/STAT and TGFB signaling pathways. Gene Rep. 2021, 22, 101012. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.R.; Park, E.G.; Kang, K.W.; Lee, S.M.; Kim, B.; Kim, H.S. Expression Analyses of MicroRNAs in Hamster Lung Tissues Infected by SARS-CoV-2. Mol. Cells 2020, 43, 953–963. [Google Scholar] [CrossRef]
- Marchi, R.; Sugita, B.; Centa, A.; Fonseca, A.S.; Bortoletto, S.; Fiorentin, K.; Ferreira, S.; Cavalli, L.R. The role of microRNAs in modulating SARS-CoV-2 infection in human cells: A systematic review. Infect. Genet. Evol. 2021, 91, 104832. [Google Scholar] [CrossRef]
- Tang, H.; Gao, Y.; Li, Z.; Miao, Y.; Huang, Z.; Liu, X.; Xie, L.; Li, H.; Wen, W.; Zheng, Y.; et al. The noncoding and coding transcriptional landscape of the peripheral immune response in patients with COVID-19. Clin. Transl. Med. 2020, 10, e200. [Google Scholar] [CrossRef]
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M.G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: The Major Cell Entry Receptor for SARS-CoV-2. Lung 2020, 198, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Chatterjee, S.; Xiao, K.; Riedel, I.; Wang, Y.; Foo, R.; Bar, C.; Thum, T. MicroRNAs targeting the SARS-CoV-2 entry receptor ACE2 in cardiomyocytes. J. Mol. Cell Cardiol. 2020, 148, 46–49. [Google Scholar] [CrossRef]
- Matarese, A.; Gambardella, J.; Sardu, C.; Santulli, G. miR-98 Regulates TMPRSS2 Expression in Human Endothelial Cells: Key Implications for COVID-19. Biomedicines 2020, 8, 462. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.W.; Zhang, Z.Y.; Song, J.; Qian, W.Z.; Gu, X.Q.; Yang, C.Y.; Shen, N.; Xue, F.; Tang, Y.J. SARS-CoV-2-Encoded MiRNAs Inhibit Host Type I Interferon Pathway and Mediate Allelic Differential Expression of Susceptible Gene. Front. Immunol. 2021, 12, 767726. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Chazal, M.; Quarato, P.; Bourdon, L.; Malabat, C.; Vallet, T.; Vignuzzi, M.; van der Werf, S.; Behillil, S.; Donati, F.; et al. A virus-derived microRNA targets immune response genes during SARS-CoV-2 infection. EMBO Rep. 2022, 23, e54341. [Google Scholar] [CrossRef] [PubMed]
- Centa, A.; Fonseca, A.S.; Ferreira, S.G.D.; Azevedo, M.L.V.; de Paula, C.B.V.; Nagashima, S.; Machado-Souza, C.; Miggiolaro, A.F.R.D.; Baena, C.P.; de Noronha, L.; et al. Deregulated miRNA expression is associated with endothelial dysfunction in post-mortem lung biopsies of COVID-19 patients. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 320, L405–L412. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.L.; Kok, K.H.; Ng, M.H.J.; Poon, V.K.M.; Yuen, K.Y.; Zheng, B.J.; Jin, D.Y. Severe Acute Respiratory Syndrome Coronavirus M Protein Inhibits Type I Interferon Production by Impeding the Formation of TRAF3 center dot TANK center dot TBK1/IKK epsilon Complex. J. Biol. Chem. 2009, 284, 16202–16209. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Li, W.; Gao, T.; Cui, Y.; Jin, Y.W.; Li, P.; Ma, Q.J.; Liu, X.; Cao, C. The Severe Acute Respiratory Syndrome Coronavirus Nucleocapsid Inhibits Type I Interferon Production by Interfering with TRIM25-Mediated RIG-I Ubiquitination. J. Virol. 2017, 91, e02143-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopecky-Bromberg, S.A.; Martinez-Sobrido, L.; Frieman, M.; Baric, R.A.; Palese, P. Severe acute respiratory syndrome coronavirus open reading frame (ORF) 3b, ORF 6, and nucleocapsid proteins function as interferon antagonists. J. Virol. 2007, 81, 548–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratia, K.; Saikatendu, K.S.; Santarsiero, B.D.; Barretto, N.; Baker, S.C.; Stevens, R.C.; Mesecar, A.D. Severe acute respiratory syndrome coronavirus papain-like protease: Structure of a viral deubiquitinating enzyme. Proc. Natl. Acad. Sci. USA 2006, 103, 5717–5722. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Xing, Y.L.; Chen, X.J.; Zheng, Y.; Yang, Y.D.; Nichols, D.B.; Clementz, M.A.; Banach, B.S.; Li, K.; Baker, S.C.; et al. Coronavirus Papain-like Proteases Negatively Regulate Antiviral Innate Immune Response through Disruption of STING-Mediated Signaling. PLoS ONE 2012, 7, e30802. [Google Scholar] [CrossRef] [Green Version]
- Bouvet, M.; Debarnot, C.; Imbert, I.; Selisko, B.; Snijder, E.J.; Canard, B.; Decroly, E. In Vitro Reconstitution of SARS-Coronavirus mRNA Cap Methylation. PLoS Pathog. 2010, 6, e1000863. [Google Scholar] [CrossRef]
- Chen, Y.; Cai, H.; Pan, J.; Xiang, N.; Tien, P.; Ahola, T.; Guo, D.Y. Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase. Proc. Natl. Acad. Sci. USA 2009, 106, 3484–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minakshi, R.; Padhan, K.; Rani, M.; Khan, N.; Ahmad, F.; Jameel, S. The SARS Coronavirus 3a Protein Causes Endoplasmic Reticulum Stress and Induces Ligand-Independent Downregulation of the Type 1 Interferon Receptor. PLoS ONE 2009, 4, e8342. [Google Scholar] [CrossRef]
- Frieman, M.; Yount, B.; Heise, M.; Kopecky-Bromberg, S.A.; Palese, P.; Baric, R.S. Severe acute respiratory syndrome coronavirus ORF6 antagonizes STAT1 function by sequestering nuclear import factors on the rough endoplasmic Reticulum/Golgi membrane. J. Virol. 2007, 81, 9812–9824. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Zhuang, M.W.; Han, L.L.; Zhang, J.; Nan, M.L.; Zhan, P.; Kang, D.W.; Liu, X.Y.; Gao, C.J.; Wang, P.H. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) membrane (M) protein inhibits type I and III interferon production by targeting RIG-I/MDA-5 signaling. Signal Transduct. Target Ther. 2020, 5, 299. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Shin, O.S. SARS-CoV-2 Nucleocapsid Protein Targets RIG-I-Like Receptor Pathways to Inhibit the Induction of Interferon Response. Cells 2021, 10, 530. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.K.; Blanco, M.R.; Bruce, E.A.; Honson, D.D.; Chen, L.M.; Chow, A.; Bhat, P.; Ollikainen, N.; Quinodoz, S.A.; Loney, C.; et al. SARS-CoV-2 Disrupts Splicing, Translation, and Protein Trafficking to Suppress Host Defenses. Cell 2020, 183, 1325–1339. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, C.; Rao, Y.; Ngo, C.; Feng, J.J.; Zhao, J.; Zhang, S.; Wang, T.Y.; Carriere, J.; Savas, A.C.; et al. SARS-CoV-2 Nsp5 Demonstrates Two Distinct Mechanisms Targeting RIG-I and MAVS To Evade the Innate Immune Response. mBio 2021, 12, e0233521. [Google Scholar] [CrossRef]
- Xia, H.J.; Cao, Z.G.; Xie, X.P.; Zhang, X.W.; Chen, J.Y.C.; Wang, H.L.; Menachery, V.D.; Rajsbaum, R.; Shi, P.Y. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020, 33, 108234. [Google Scholar] [CrossRef]
- Wang, W.J.; Zhou, Z.; Xiao, X.; Tian, Z.Q.; Dong, X.J.; Wang, C.H.; Li, L.; Ren, L.L.; Lei, X.B.; Xiang, Z.C.; et al. SARS-CoV-2 nsp12 attenuates type I interferon production by inhibiting IRF3 nuclear translocation. Cell. Mol. Immunol. 2021, 18, 945–953. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.W.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.F.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Hsu, J.C.C.; Laurent-Rolle, M.; Pawlak, J.B.; Wilen, C.B.; Cresswell, P. Translational shutdown and evasion of the innate immune response by SARS-CoV-2 NSP14 protein. Proc. Natl. Acad. Sci. USA 2021, 118, e2101161118. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.J.; Su, J.M.; Shen, S.; Hu, Y.; Huang, D.B.; Zheng, W.W.; Lou, M.; Shi, Y.F.; Wang, M.; Chen, S.Q.; et al. Unique and complementary suppression of cGAS-STING and RNA sensing- triggered innate immune responses by SARS-CoV-2 proteins. Signal Transduct. Target Ther. 2021, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.B.; Dong, X.J.; Ma, R.Y.; Wang, W.J.; Xiao, X.; Tian, Z.Q.; Wang, C.H.; Wang, Y.; Li, L.; Ren, L.L.; et al. Activation and evasion of type I interferon responses by SARS-CoV-2. Nat. Commun. 2020, 11, 3810. [Google Scholar] [CrossRef] [PubMed]
- Kouwaki, T.; Nishimura, T.; Wang, G.; Oshiumi, H. RIG-I-Like Receptor-Mediated Recognition of Viral Genomic RNA of Severe Acute Respiratory Syndrome Coronavirus-2 and Viral Escape from the Host Innate Immune Responses. Front. Immunol. 2021, 12, 700926. [Google Scholar] [CrossRef]
- Rashid, F.; Dzakah, E.E.; Wang, H.; Tang, S. The ORF8 protein of SARS-CoV-2 induced endoplasmic reticulum stress and mediated immune evasion by antagonizing production of interferon beta. Virus Res. 2021, 296, 198350. [Google Scholar] [CrossRef]
- Han, L.L.; Zhuang, M.W.; Deng, J.; Zheng, Y.; Zhang, J.; Nan, M.L.; Zhang, X.J.; Gao, C.J.; Wang, P.H. SARS-CoV-2 ORF9b antagonizes type I and III interferons by targeting multiple components of the RIG-I/MDA-5-MAVS, TLR3-TRIF, and cGAS-STING signaling pathways. J. Med. Virol. 2021, 93, 5376–5389. [Google Scholar] [CrossRef]
- Thorne, L.G.; Bouhaddou, M.; Reuschl, A.K.; Zuliani-Alvarez, L.; Polacco, B.; Pelin, A.; Batra, J.; Whelan, M.V.X.; Hosmillo, M.; Fossati, A.; et al. Evolution of enhanced innate immune evasion by SARS-CoV-2. Nature 2022, 602, 487–495. [Google Scholar] [CrossRef]
- Wu, J.; Shi, Y.H.; Pan, X.Y.; Wu, S.; Hou, R.X.; Zhang, Y.; Zhong, T.S.; Tang, H.; Du, W.; Wang, L.Y.; et al. SARS-CoV-2 ORF9b inhibits RIG-I-MAVS antiviral signaling by interrupting K63-linked ubiquitination of NEMO. Cell Rep. 2021, 34, 108761. [Google Scholar] [CrossRef]
- Li, X.; Hou, P.; Ma, W.; Wang, X.; Wang, H.; Yu, Z.; Chang, H.; Wang, T.; Jin, S.; Wang, X.; et al. SARS-CoV-2 ORF10 suppresses the antiviral innate immune response by degrading MAVS through mitophagy. Cell Mol. Immunol. 2022, 19, 67–78. [Google Scholar] [CrossRef]
- Comar, C.E.; Goldstein, S.A.; Li, Y.Z.; Yount, B.; Baric, R.S.; Weiss, S.R. Antagonism of dsRNA-Induced Innate Immune Pathways by NS4a and NS4b Accessory Proteins during MERS Coronavirus Infection. mBio 2019, 10, e00319-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornbrough, J.M.; Jha, B.K.; Yount, B.; Goldstein, S.A.; Li, Y.Z.; Elliott, R.; Sims, A.C.; Baric, R.S.; Silverman, R.H.; Weiss, S.R. Middle East Respiratory Syndrome Coronavirus NS4b Protein Inhibits Host RNase L Activation. mBio 2016, 7, e00258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canton, J.; Fehr, A.R.; Fernandez-Delgado, R.; Gutierrez-Alvarez, F.J.; Sanchez-Aparicio, M.T.; García-Sastre, A.; Perlman, S.; Enjuanes, L.; Sola, I. MERS-CoV 4b protein interferes with the NF-κB-dependent innate immune response during infection. PLoS Pathog. 2018, 14, e1006838. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, D.; Zillinger, T.; Muth, D.; Zielecki, F.; Horvath, G.; Suliman, T.; Barchet, W.; Weber, F.; Drosten, C.; Muller, M.A. Middle East Respiratory Syndrome Coronavirus Accessory Protein 4a Is a Type I Interferon Antagonist. J. Virol. 2013, 87, 12489–12495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, K.L.; Yeung, M.L.; Kok, K.H.; Yuen, K.S.; Kew, C.; Lui, P.Y.; Chan, C.P.; Tse, H.; Woo, P.C.Y.; Yuen, K.Y.; et al. Middle East Respiratory Syndrome Coronavirus 4a Protein Is a Double-Stranded RNA-Binding Protein That Suppresses PACT-Induced Activation of RIG-I and MDA5 in the Innate Antiviral Response. J. Virol. 2014, 88, 4866–4876. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ye, F.; Zhu, N.; Wang, W.L.; Deng, Y.; Zhao, Z.D.; Tan, W.J. Middle East respiratory syndrome coronavirus ORF4b protein inhibits type I interferon production through both cytoplasmic and nuclear targets. Sci. Rep. 2015, 5, 17554. [Google Scholar] [CrossRef] [Green Version]
- Menachery, V.D.; Mitchell, H.D.; Cockrell, A.S.; Gralinski, L.E.; Yount, B.L.; Graham, R.L.; McAnarney, E.T.; Douglas, M.G.; Scobey, T.; Beall, A.; et al. MERS-CoV Accessory ORFs Play Key Role for Infection and Pathogenesis. mBio 2017, 8, e00665-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liao, Y.; Yap, P.L.; Png, K.J.; Tam, J.P.; Liu, D.X. Inhibition of protein kinase R activation and upregulation of GADD34 expression play a synergistic role in facilitating coronavirus replication by maintaining de novo protein synthesis in virus-infected cells. J. Virol. 2009, 83, 12462–12472. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Shaw, N.; Yan, L.; Lou, Z.; Rao, Z. Structural view and substrate specificity of papain-like protease from avian infectious bronchitis virus. J. Biol. Chem. 2015, 290, 7160–7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kint, J.; Langereis, M.A.; Maier, H.J.; Britton, P.; van Kuppeveld, F.J.; Koumans, J.; Wiegertjes, G.F.; Forlenza, M. Infectious Bronchitis Coronavirus Limits Interferon Production by Inducing a Host Shutoff That Requires Accessory Protein 5b. J. Virol. 2016, 90, 7519–7528. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ke, H.; Blikslager, A.; Fujita, T.; Yoo, D. Type III Interferon Restriction by Porcine Epidemic Diarrhea Virus and the Role of Viral Protein nsp1 in IRF1 Signaling. J. Virol. 2018, 92, e01677-17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Shi, K.; Yoo, D. Suppression of type I interferon production by porcine epidemic diarrhea virus and degradation of CREB-binding protein by nsp1. Virology 2016, 489, 252–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Ma, J.; Yoo, D. Inhibition of NF-κB activity by the porcine epidemic diarrhea virus nonstructural protein 1 for innate immune evasion. Virology 2017, 510, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Cheng, L.; Xu, J.; Li, P.; Li, X.; Zou, D.; Zhang, Y.; Wang, X.; Wu, X.; Shen, Y.; et al. The accessory protein ORF3 of porcine epidemic diarrhea virus inhibits cellular interleukin-6 and interleukin-8 productions by blocking the nuclear factor-κB p65 activation. Vet. Microbiol. 2020, 251, 108892. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Chen, J.; Tu, J.; Zhang, B.; Chen, X.; Shi, H.; Baker, S.C.; Feng, L.; Chen, Z. The papain-like protease of porcine epidemic diarrhea virus negatively regulates type I interferon pathway by acting as a viral deubiquitinase. J. Gen. Virol. 2013, 94 Pt 7, 1554–1567. [Google Scholar] [CrossRef]
- Wang, D.; Fang, L.; Shi, Y.; Zhang, H.; Gao, L.; Peng, G.; Chen, H.; Li, K.; Xiao, S. Porcine Epidemic Diarrhea Virus 3C-Like Protease Regulates Its Interferon Antagonism by Cleaving NEMO. J. Virol. 2016, 90, 2090–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhang, H.; Shi, Z.; Chen, J.; Li, M.; Shi, H.; Shi, D.; Guo, L.; Feng, L. Porcine Epidemic Diarrhea Virus nsp15 Antagonizes Interferon Signaling by RNA Degradation of TBK1 and IRF3. Viruses 2020, 12, 599. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Su, Y.; Li, R.; Liang, Z.; Dong, S.; Huang, J. PEDV nsp16 negatively regulates innate immunity to promote viral proliferation. Virus Res. 2019, 265, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Liu, Z.Q.; Li, G.W.; Chen, C.; Luo, H.; Liu, Y.J.; Zhuo, X.H.; Shi, X.F.; Fang, W.H.; Li, X.L. Nucleocapsid protein from porcine epidemic diarrhea virus isolates can antagonize interferon-λ production by blocking the nuclear factor-κB nuclear translocation. J. Zhejiang Univ. Sci. B 2018, 19, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Fang, L.; Jing, H.; Zeng, S.; Wang, D.; Liu, L.; Zhang, H.; Luo, R.; Chen, H.; Xiao, S. Porcine epidemic diarrhea virus nucleocapsid protein antagonizes beta interferon production by sequestering the interaction between IRF3 and TBK1. J. Virol. 2014, 88, 8936–8945. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Wang, X.; Guo, D.; Cao, J.; Cheng, L.; Li, X.; Zou, D.; Zhang, Y.; Xu, J.; Wu, X.; et al. Porcine epidemic diarrhea virus E protein suppresses RIG-I signaling-mediated interferon-β production. Vet. Microbiol. 2021, 254, 108994. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, A.; Sun, Y.; Zhang, S.; Xia, T.; Guo, T.; Hao, Z.; Sun, L.; Jiang, Y.; Qiao, X.; et al. Porcine transmissible gastroenteritis virus inhibits NF-κB activity via nonstructural protein 3 to evade host immune system. Virol. J. 2019, 16, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindler, E.; Thiel, V.; Weber, F. Interaction of SARS and MERS Coronaviruses with the Antiviral Interferon Response. Adv. Virus Res. 2016, 96, 219–243. [Google Scholar] [PubMed]
- Costa, T.J.; Potje, S.R.; Fraga-Silva, T.F.C.; da Silva-Neto, J.A.; Barros, P.R.; Rodrigues, D.; Machado, M.R.; Martins, R.B.; Santos-Eichler, R.A.; Benatti, M.N.; et al. Mitochondrial DNA and TLR9 activation contribute to SARS-CoV-2-induced endothelial cell damage. Vasc. Pharmacol. 2022, 142, 106946. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.M.; Zhang, J.; Li, P.W.; Hu, Y.W.; Cao, L.; Ouyang, S.; Bi, Y.H.; Nie, P.; Chang, M.X. NOD1 Promotes Antiviral Signaling by Binding Viral RNA and Regulating the Interaction of MDA5 and MAVS. J. Immunol. 2020, 204, 2216–2231. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram of innate immune response caused by IAV RNA. Upon IAV infection, viral ssRNA in the endosome is sensed by TLR7/8 and recruits the MyD88 adapter. Activated MyD88 induces the production of inflammatory cytokines through the TAK1-IKK-NF-κB signaling pathway. Viral double-stranded RNA in the endosome is sensed by TLR3, which further recruits TRIF and PI3K adapters, inducing the production of IFN through the TAK1-IRFs signaling pathway. Viral dsRNA in the cytoplasm is sensed by RIG-I and IFI16, leading to activation of NF-κB and phosphorylation of IRFs by binding to MAVS on mitochondria. Furthermore, IAV RNA can be sensed by NLRP3, which promotes the formation of inflammasomes, causing pyroptosis and secreting the IL-1β and IL-18. Of note, there is no direct evidence to demonstrate that IAV RNA can be recognized by DDX3X and MDA5 in mammals, which is indicated by dotted lines. Abbreviation: ROS, reactive oxygen species; MLKL, mixed lineage kinase domain-like pseudokinase. Other abbreviations are shown in the legend to Figure 2.
Figure 1.
Schematic diagram of innate immune response caused by IAV RNA. Upon IAV infection, viral ssRNA in the endosome is sensed by TLR7/8 and recruits the MyD88 adapter. Activated MyD88 induces the production of inflammatory cytokines through the TAK1-IKK-NF-κB signaling pathway. Viral double-stranded RNA in the endosome is sensed by TLR3, which further recruits TRIF and PI3K adapters, inducing the production of IFN through the TAK1-IRFs signaling pathway. Viral dsRNA in the cytoplasm is sensed by RIG-I and IFI16, leading to activation of NF-κB and phosphorylation of IRFs by binding to MAVS on mitochondria. Furthermore, IAV RNA can be sensed by NLRP3, which promotes the formation of inflammasomes, causing pyroptosis and secreting the IL-1β and IL-18. Of note, there is no direct evidence to demonstrate that IAV RNA can be recognized by DDX3X and MDA5 in mammals, which is indicated by dotted lines. Abbreviation: ROS, reactive oxygen species; MLKL, mixed lineage kinase domain-like pseudokinase. Other abbreviations are shown in the legend to Figure 2.

Figure 2.
Schematic diagram of signaling transduction pathways in host response induced by recognition of PAMPs by PRRs. Sensing PAMPs, the PRRs recruit and activate MyD88, TRIF, TIRAP, and other adapters. These adapters transduce activating signals, which finally induce activation of transcriptional regulators NF-κB and IRFs, leading to the expression of IFNs and proinflammatory cytokines or mediators, such as IL-1, IL-6, IL-12, TNF-α, NO, etc. Abbreviation: MyD88, myeloid differentiation factor 88; TRIF, TIR domain-containing adapter-inducing IFN-β; TIRAP, TIR domain-containing adapter protein; IRAK, IL-1 receptor kinase; TRAF, tumor necrosis factor receptor-associated factor; MALT1, mucosa-associated lymphoid tissue lymphoma translocation gene 1; Syk, spleen tyrosine kinase; TAK, TGF-β activated kinase; NEMO, NF-κB essential modulator; IKK, inhibitor of NF-κB kinase; IκB, inhibitor of NF-κB; MAVS, mitochondrial antiviral signaling protein; PI3K, phosphoinositol-3 kinase; NF-κB, nuclear factor kappa enhancer binding protein; IRF, interferon regulatory factor; RIG-I, retinoic acid-inducible gene I; MDA5, melanoma differentiation-associated gene 5; STING, stimulator of interferon genes; LPS, Lipopolysaccharide; GSDMD, gasdermin D.
Figure 2.
Schematic diagram of signaling transduction pathways in host response induced by recognition of PAMPs by PRRs. Sensing PAMPs, the PRRs recruit and activate MyD88, TRIF, TIRAP, and other adapters. These adapters transduce activating signals, which finally induce activation of transcriptional regulators NF-κB and IRFs, leading to the expression of IFNs and proinflammatory cytokines or mediators, such as IL-1, IL-6, IL-12, TNF-α, NO, etc. Abbreviation: MyD88, myeloid differentiation factor 88; TRIF, TIR domain-containing adapter-inducing IFN-β; TIRAP, TIR domain-containing adapter protein; IRAK, IL-1 receptor kinase; TRAF, tumor necrosis factor receptor-associated factor; MALT1, mucosa-associated lymphoid tissue lymphoma translocation gene 1; Syk, spleen tyrosine kinase; TAK, TGF-β activated kinase; NEMO, NF-κB essential modulator; IKK, inhibitor of NF-κB kinase; IκB, inhibitor of NF-κB; MAVS, mitochondrial antiviral signaling protein; PI3K, phosphoinositol-3 kinase; NF-κB, nuclear factor kappa enhancer binding protein; IRF, interferon regulatory factor; RIG-I, retinoic acid-inducible gene I; MDA5, melanoma differentiation-associated gene 5; STING, stimulator of interferon genes; LPS, Lipopolysaccharide; GSDMD, gasdermin D.

Figure 3.
Schematic diagram of innate immune response induced by coronavirus RNA. Viral dsRNA is sensed by TLR3 in endosomes, triggering the innate immune response, while ssRNA in endosomes is sensed by TLR7/8 and induces the production of inflammatory cytokines through the MyD88-TAK1-IKK-NF-κB signaling pathway. Viral dsRNA is recognized by MDA5 and NOD1, and then interacts with MAVS on mitochondria, which further activates the NF-κB and IRFs signaling pathways. Viral GU-rich ssRNA is sensed by TLR8, causing the activation of NLRP3 inflammasomes through the Caspase-8-RIPK1 signaling pathway, leading to the secretion of mature IL-1β and IL-18. Importantly, it is not clear whether viral dsRNA is recognized by RIG-I to initiate IFN response or whether viral RNA induces inflammasome formation; thus, the associated pathway is indicated by dotted lines. Abbreviation: RIPK, Receptor interacting protein kinase; other abbreviations are shown in the legend to Figure 2.
Figure 3.
Schematic diagram of innate immune response induced by coronavirus RNA. Viral dsRNA is sensed by TLR3 in endosomes, triggering the innate immune response, while ssRNA in endosomes is sensed by TLR7/8 and induces the production of inflammatory cytokines through the MyD88-TAK1-IKK-NF-κB signaling pathway. Viral dsRNA is recognized by MDA5 and NOD1, and then interacts with MAVS on mitochondria, which further activates the NF-κB and IRFs signaling pathways. Viral GU-rich ssRNA is sensed by TLR8, causing the activation of NLRP3 inflammasomes through the Caspase-8-RIPK1 signaling pathway, leading to the secretion of mature IL-1β and IL-18. Importantly, it is not clear whether viral dsRNA is recognized by RIG-I to initiate IFN response or whether viral RNA induces inflammasome formation; thus, the associated pathway is indicated by dotted lines. Abbreviation: RIPK, Receptor interacting protein kinase; other abbreviations are shown in the legend to Figure 2.

Table 2.
The comparison of innate immune sensors involved in recognition of MERS-CoV, SARS-CoV, and SARS-CoV-2 infections.
Table 2.
The comparison of innate immune sensors involved in recognition of MERS-CoV, SARS-CoV, and SARS-CoV-2 infections.
| Virus | TLRs | RLRs | NLRs | CLR | Others |
|---|---|---|---|---|---|
| MERS-CoV | TLR7 [190] TLR8 [191] | RIG-I [211] MDA5 [211] | NLRP3 [212] | Mincle [211] | PKR [213] |
| SARS-CoV | TLR7 [189] TLR8 [191] TLR4 [183] TLR2 [214] | N/A 1 | NLRP3 [215] | N/A | N/A |
| SARS-CoV-2 | TLR3 [184] TLR7 [184] TLR8 [191] TLR4 [216] TLR2 [201] | MDA5 [196] LGP2 [196] RIG-I [201] | NOD1 [196] NLRP3 [207] | N/A | PKR [217] OAS [217] |
1 N/A: not applicable (data for the corresponding sensor has not been reported).
Table 3.
The roles of coronavirus proteins in innate immune response across the species.
| Host | Virus | Viral Protein | Protein Function | References |
|---|---|---|---|---|
| Human | SARS-CoV | M | Delay the formation of TRAF3-containing complex | [245] |
| N | Inhibit the ubiquitination of RIG-I mediated by TRIM25 | [246] | ||
| N, ORF3b, ORF6 | Inhibit the activation of IRF3 and the translocation of NF-κB | [247] | ||
| NSP1 | Decrease the phosphorylation of STAT1 | [178] | ||
| PLpro(Nsp3) | Inhibit the ubiquitination of STING, TBK1 and IRF3 | [248,249] | ||
| Nsp14, Nsp16, Nsp10 | Induce viral mRNA cap methylation and escape the detection of MDA5 | [250,251] | ||
| ORF3a | Degrade the expression of IFNAR1 | [252] | ||
| ORF6 | Inhibit the translocation of STAT1 | [253] | ||
| SARS-CoV-2 | M | Degrade the ubiquitinated TBK1 and inhibit IFN-I response | [254] | |
| N | Interact with RIG-I and suppress the RIG-I signaling pathway | [255] | ||
| Nsp1 | Bind to mRNA entry channel of the ribosome and inhibit ISGs mRNA translation | [256] | ||
| Nsp5 | Cleave RIG-I and promote the proteosome-mediated degradation of MAVS | [257] | ||
| Nsp6 | Inhibit the activation of IRF3 and suppress the phosphorylation of STAT1 and STAT2 | [258] | ||
| Nsp8, Nsp9 | Disrupt the function of signal recognition particle (SRP) complex and suppress IFN trafficking | [256] | ||
| Nsp16 | Inhibit ISGs mRNA splicing and suppress innate immune response | [256] | ||
| Nsp12 | Inhibit the IRF3 nuclear translocation and attenuate IFN-I production | [259] | ||
| Nsp13 | Interact with TBK1 and inhibit its activation | [260] | ||
| Nsp14 | Shut down the protein synthesis and abolish the production of ISGs | [261] | ||
| Nsp15 | Interact with RNF41 and inhibit the activation of TBK1 | [260] | ||
| ORF3a | Interact with STING and inhibit the nuclear import of NF-κB | [262] | ||
| ORF6 | Interact with NUP98-RAE1 Inhibit STAT1 nuclear translocation | [260] [263] | ||
| ORF7a | Destabilize the TBK1 and lead to attenuated IRF-3 phosphorylation | [264] | ||
| ORF8 | Induce ER stress and inhibit the nuclear translocation of IRF3 | [265] | ||
| ORF9b | Interact with RIG-I, MDA-5, and STING and inhibit the phosphorylation of IRF3 | [266] | ||
| Interact with TOM70 and suppress the innate immune response | [267] | |||
| Interrupt the K63-linked polyubiquitination of NEMO and inhibit IKKα/β/γ-NF-κB signaling pathway | [268] | |||
| ORF10 | Degrade MAVS through mitophagy | [269] | ||
| MERS-CoV | NS4a | Impede the activation of PKR | [213,270] | |
| NS4b | Inhibit the activation of RNaseL | [271] | ||
| Bind to karyopherin-α4 (KPNA4) and inhibit the translocate of NF-κB | [272] | |||
| ORF4a | Inhibit the activation of MDA5 | [273,274] | ||
| ORF4b | Inhibit the formation of TBK1 signaling complex | [275] | ||
| ORF5 | Inhibit the activation of NF-κB | [276] | ||
| Chicken | IBV | Nsp2 | Inhibit the activation of PKR | [277] |
| Nsp3 | Interrupt the activation of RIG-I and NF-κB | [278] | ||
| Accessory protein 5b | Induce a host shutoff and suppress IFN production | [279] | ||
| Swine | PEDV | Nsp1 | Inhibit the activation of IRF1/IRF3 | [280] |
| Degrade CREB-binding protein and inhibit IFN-I production | [281] | |||
| Inhibit the translocation of NF-κB | [282] | |||
| ORF3 | Inhibit the phosphorylation of IκBα and NF-κB translocation | [283] | ||
| PLP (Nsp3) | Deubiquitinate RIG-I and STING | [284] | ||
| 3CLpro (Nsp5) | Cleave NEMO to inhibit the innate immune response | [285] | ||
| Nsp15 | RNA degradation of TBK1 and IRF3. | [286] | ||
| Nsp16 | Inhibit the activation of RIG-I and MDA5 | [287] | ||
| N | Inhibit the translocation of NF-κB | [288] | ||
| Interrupt the interaction between IRF3 and TBK1 | [289] | |||
| E | Inhibit the RIG-I mediated IFN-β production | [290] | ||
| TGEV | Nsp3 | Inhibit the activation of NF-κB | [291] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, W.; Wang, H.; Zheng, S.J. Roles of RNA Sensors in Host Innate Response to Influenza Virus and Coronavirus Infections. Int. J. Mol. Sci. 2022, 23, 8285. https://doi.org/10.3390/ijms23158285
AMA Style
Li W, Wang H, Zheng SJ. Roles of RNA Sensors in Host Innate Response to Influenza Virus and Coronavirus Infections. International Journal of Molecular Sciences. 2022; 23(15):8285. https://doi.org/10.3390/ijms23158285
Chicago/Turabian StyleLi, Wei, Hongnuan Wang, and Shijun J. Zheng. 2022. "Roles of RNA Sensors in Host Innate Response to Influenza Virus and Coronavirus Infections" International Journal of Molecular Sciences 23, no. 15: 8285. https://doi.org/10.3390/ijms23158285
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.