
Potent Therapeutic Strategies for COVID-19 with Single-Domain Antibody Immunoliposomes Neutralizing SARS-CoV-2 and Lip/cGAMP Enhancing Protective Immunity

Abstract
:1. Introduction
2. Results and Discussion
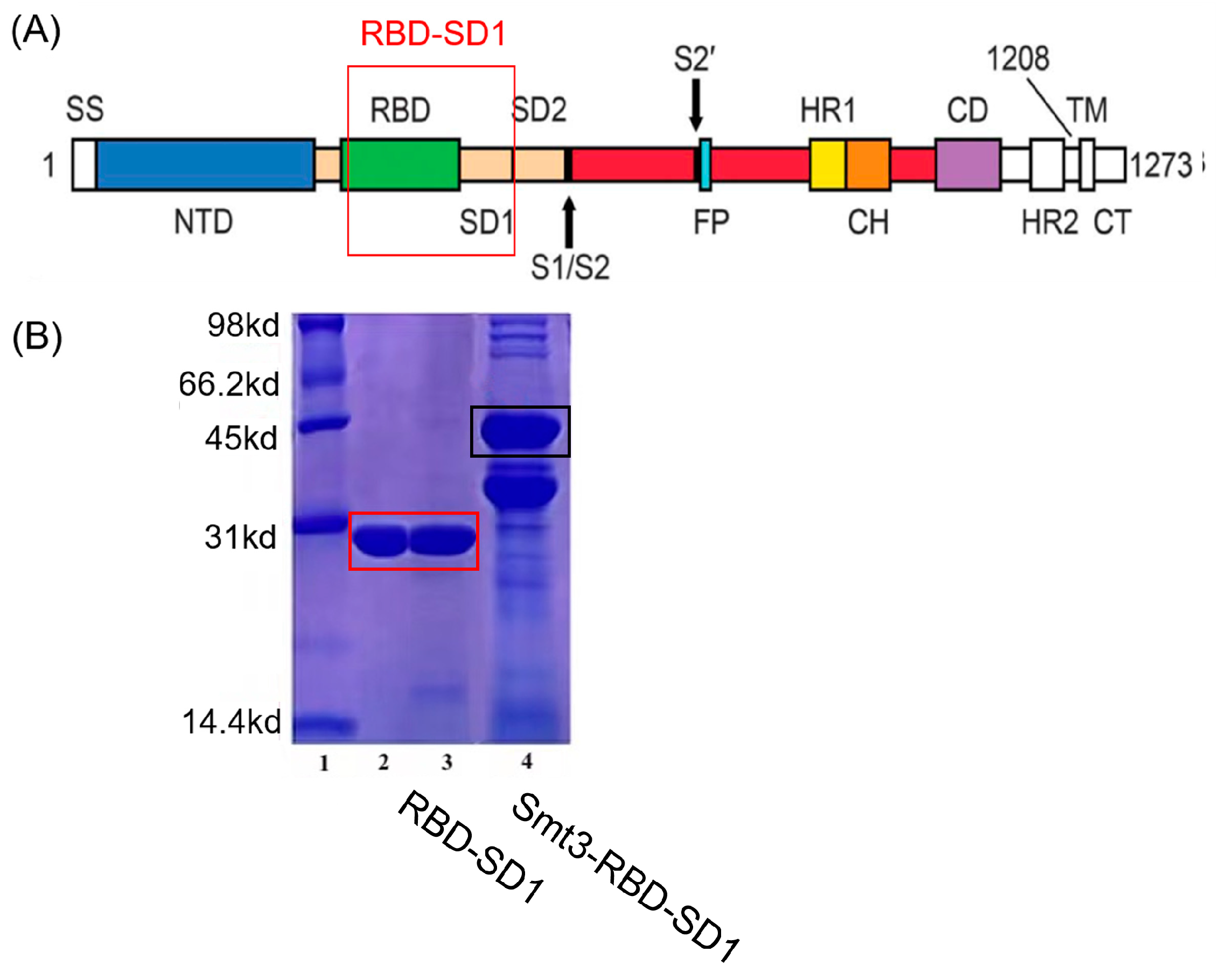
2.1. Preparation and Characterization of Nanobodies and Immunoliposomes
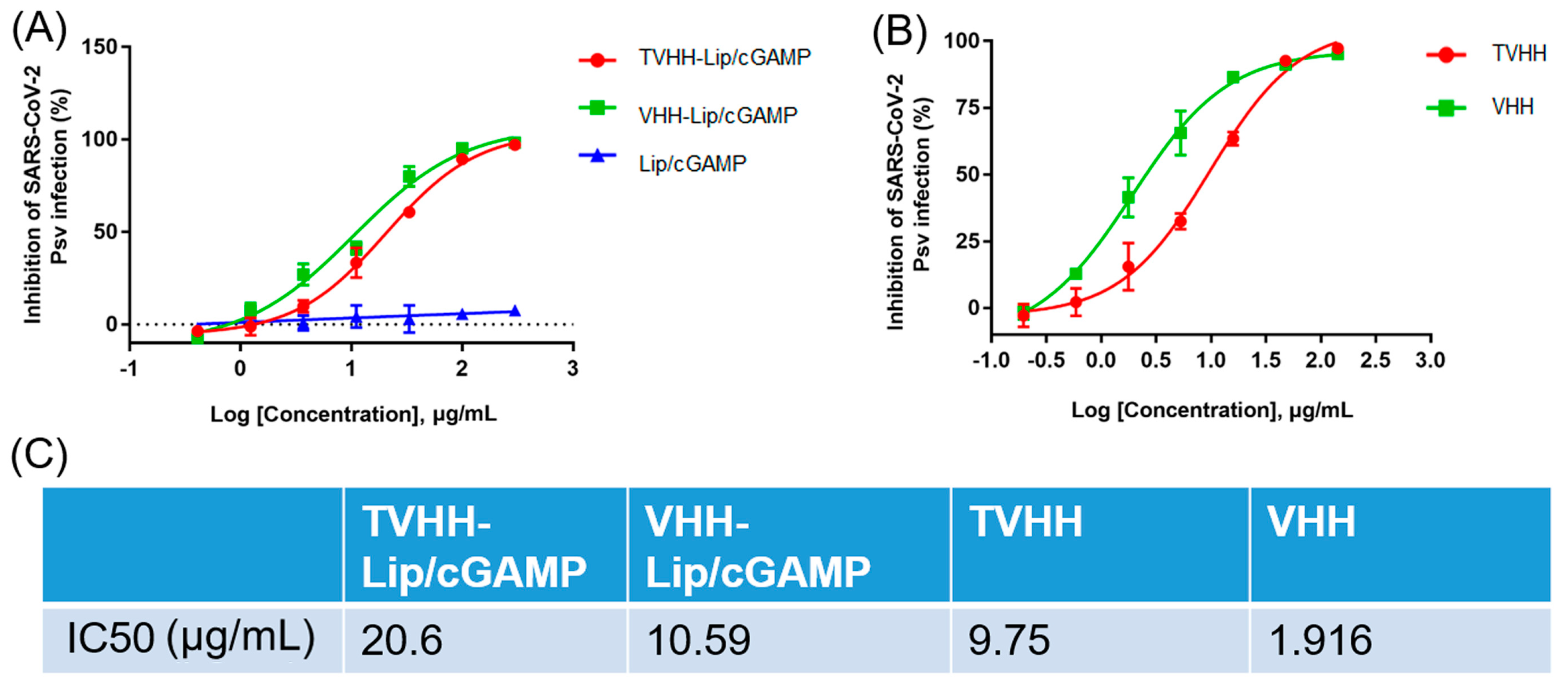
2.2. Neutralization of Nanobodies and Immunoliposomes
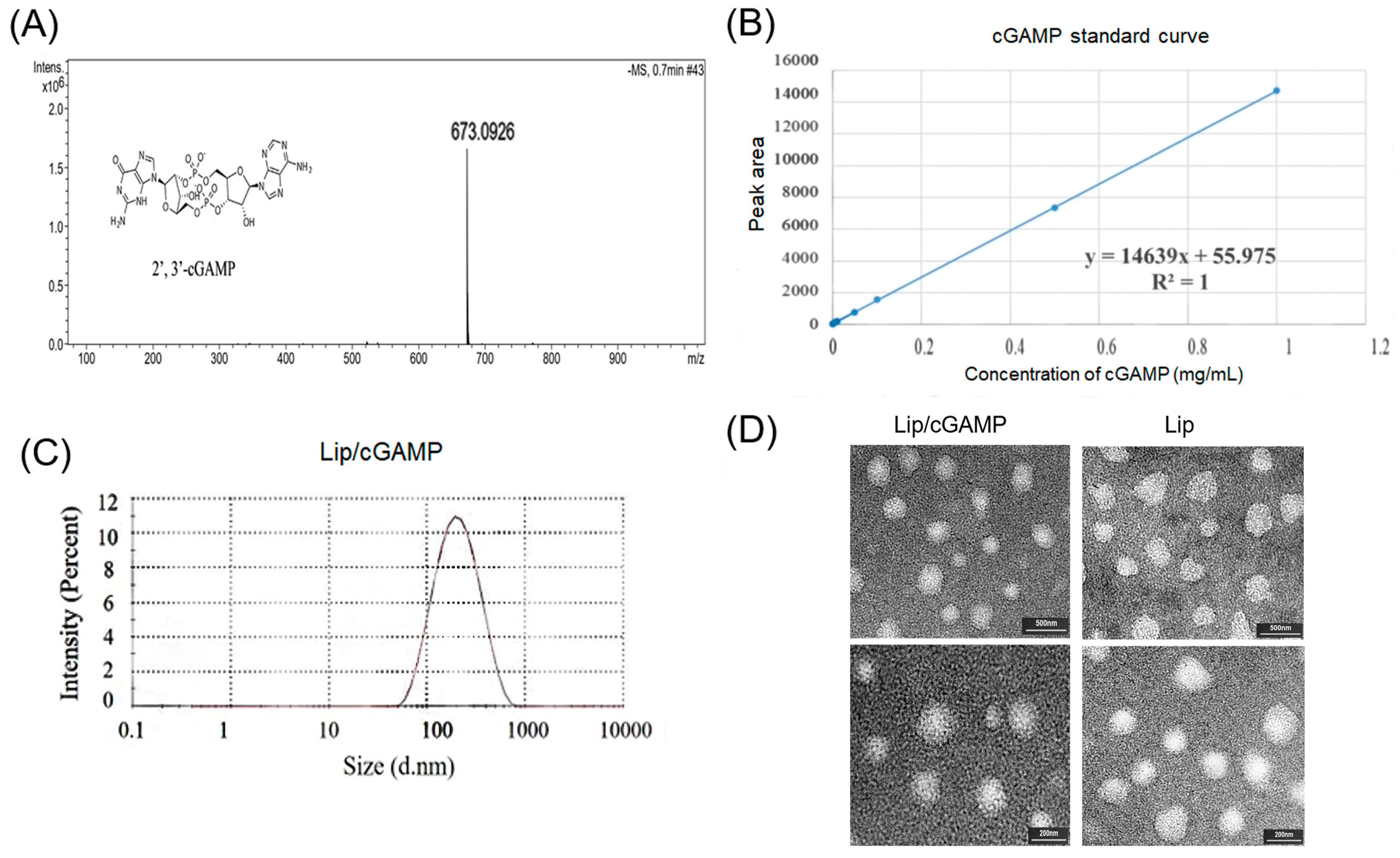
2.3. Preparation and Characterization of Antigen RBD-SD1 and Lip-cGAMP
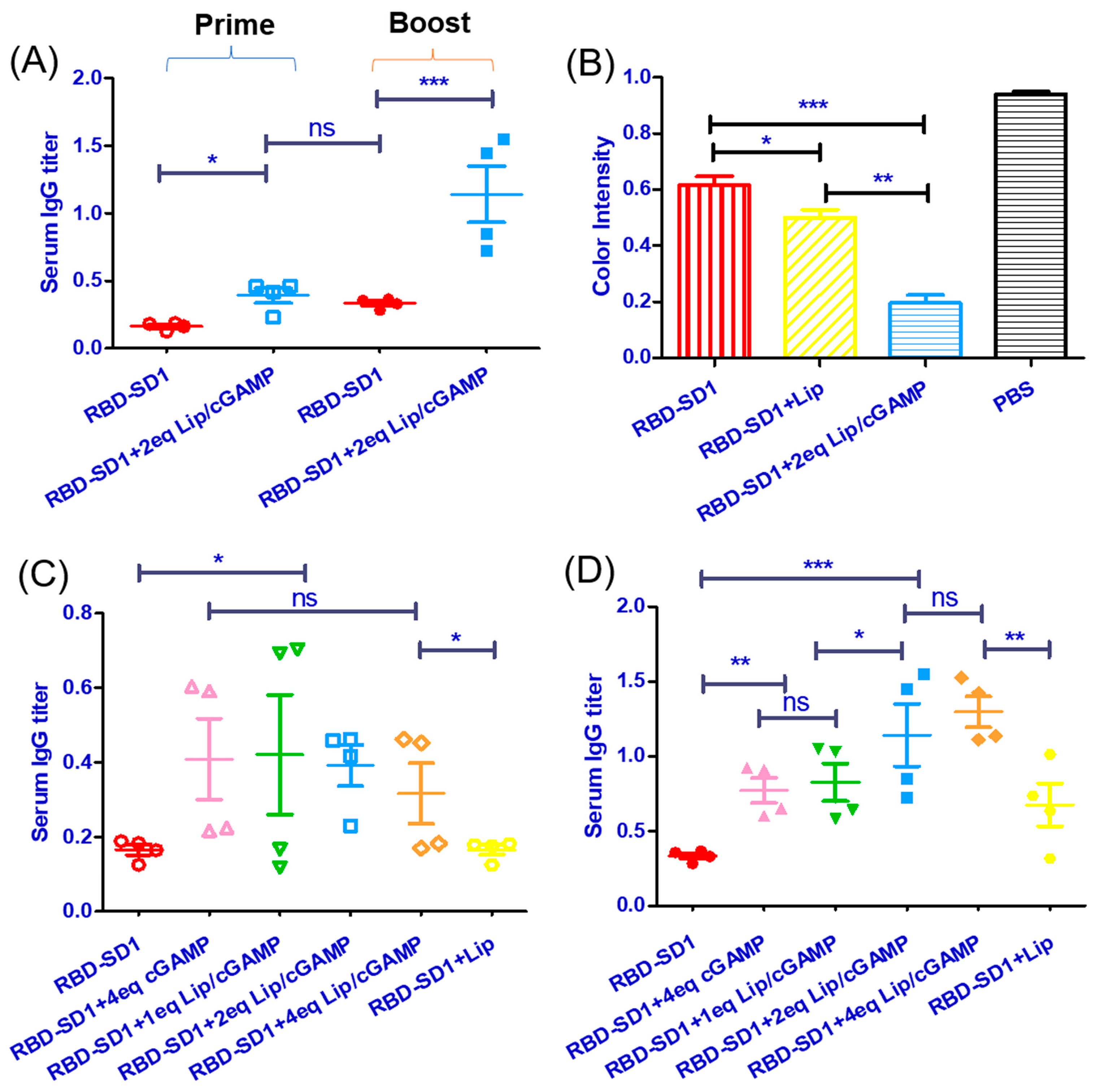
2.4. Imunization of Mice with the Recombinant Vaccine RBD-SD1 and Lip-cGAMP
3. Materials and Methods
3.1. Prepration of Nanobodies and RBD-SD1
3.2. Synthesis of Nanobody-Modified Liposomes
3.3. Characterization of cGAMP-loaded Liposomes
3.4. Pseudotyped Virus Neutralization
3.5. Enzyme-Linked Immunosorbent Assay (ELISA)
3.6. Immunize mice and Antibody Measurement
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Kissler, S.M.; Tedijanto, C.; Goldstein, E.; Grad, Y.H.; Lipsitch, M. Projecting the transmission dynamics of SARS-CoV-2 through the postpandemic period. Science 2020, 368, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.L.; Wang, Y.M.; Li, X.W.; Ren, L.L.; Zhao, J.P.; Hu, Y.; Zhang, L.; Fan, G.H.; Xu, J.Y.; Gu, X.Y.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrapp, D.; Wang, N.S.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.S.; Luo, C.M.; Ye, G.; Geng, Q.B.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Xiang, R.; Deng, X.Q.; Wang, L.L.; Yu, Z.S.; Tian, S.J.; Liang, R.Y.; Li, Y.B.; Ying, T.L.; Jiang, S.B. Receptor-binding domain-specific human neutralizing monoclonal antibodies against SARS-CoV and SARS-CoV-2. Signal Transduct. Target. Ther. 2020, 5, 212. [Google Scholar] [CrossRef]
- Wan, J.K.; Xing, S.H.; Ding, L.F.; Wang, Y.H.; Gu, C.J.; Wu, Y.L.; Rong, B.W.; Li, C.; Wang, S.Q.; Chen, K.; et al. Human-IgG-Neutralizing Monoclonal Antibodies Block the SARS-CoV-2 Infection. Cell Rep. 2020, 32, 107918. [Google Scholar] [CrossRef]
- Yang, J.Y.; Wang, W.; Chen, Z.M.; Lu, S.Y.; Yang, F.L.; Bi, Z.F.; Bao, L.L.; Mo, F.; Li, X.; Huang, Y.; et al. A vaccine targeting the RBD of the S protein of SARS-CoV-2 induces protective immunity. Nature 2020, 586, 572–577. [Google Scholar] [CrossRef]
- Zost, S.J.; Gilchuk, P.; Case, J.B.; Binshtein, E.; Chen, R.T.E.; Nkolola, J.P.; Schafer, A.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.L.; Li, C.; Huang, A.L.; Xia, S.; Lu, S.C.; Shi, Z.L.; Lu, L.; Jiang, S.B.; Yang, Z.L.; Wu, Y.L.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.H.; Wang, P.F.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.W.; Sahi, V.; Figueroa, A.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Wolfson, W. Ablynx makes nanobodies from llama bodies. Chem. Biol. 2006, 13, 1243–1244. [Google Scholar] [CrossRef] [Green Version]
- Khodabakhsh, F.; Behdani, M.; Rami, A.; Kazemi-Lomedasht, F. Single-Domain Antibodies or Nanobodies: A Class of Next-Generation Antibodies. Int. Rev. Immunol. 2018, 37, 316–322. [Google Scholar] [CrossRef]
- Wrapp, D.; De Vlieger, D.; Corbett, K.S.; Torres, G.M.; Wang, N.S.; Van Breedam, W.; Roose, K.; van Schie, L.; Hoffmann, M.; Pohlmann, S.; et al. Structural Basis for Potent Neutralization of Betacoronaviruses by Single-Domain Camelid Antibodies. Cell 2020, 181, 1004–1015. [Google Scholar] [CrossRef]
- Wu, Y.L.; Li, C.; Xia, S.; Tian, X.L.; Kong, Y.; Wang, Z.; Gu, C.J.; Zhang, R.; Tu, C.; Xie, Y.H.; et al. Identification of Human Single-Domain Antibodies against SARS-CoV-2. Cell Host Microbe 2020, 27, 891–898. [Google Scholar] [CrossRef]
- Wu, J.X.; Sun, L.J.; Chen, X.; Du, F.H.; Shi, H.P.; Chen, C.; Chen, Z.J.J. Cyclic GMP-AMP Is an Endogenous Second Messenger in Innate Immune Signaling by Cytosolic DNA. Science 2013, 339, 826–830. [Google Scholar] [CrossRef] [Green Version]
- Ablasser, A.; Goldeck, M.; Cavlar, T.; Deimling, T.; Witte, G.; Rohl, I.; Hopfner, K.P.; Ludwig, J.; Hornung, V. cGAS produces a 2’-5’-linked cyclic dinucleotide second messenger that activates STING. Nature 2013, 498, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Bai, X.C.; Chen, Z.J.J. Structures and Mechanisms in the cGAS-STING Innate Immunity Pathway. Immunity 2020, 53, 43–53. [Google Scholar] [CrossRef]
- Tao, J.L.; Zhou, X.; Jiang, Z.F. cGAS-cGAMP-STING: The Three Musketeers of Cytosolic DNA Sensing and Signaling. IUBMB Life 2016, 68, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Chen, Z.J.J. Intrinsic antiviral immunity. Nat. Immunol. 2012, 13, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.H.; Ferretti, M.; Ying, B.L.; Descamps, H.; Lee, E.; Dittmar, M.; Lee, J.S.; Whig, K.; Kamalia, B.; Dohnalova, L.; et al. Pharmacological activation of STING blocks SARS-CoV-2 infection. Sci. Immunol. 2021, 6, eabi9007. [Google Scholar] [CrossRef] [PubMed]
- Ramanjulu, J.M.; Pesiridis, G.S.; Yang, J.S.; Concha, N.; Singhaus, R.; Zhang, S.Y.; Tran, J.L.; Moore, P.; Lehmann, S.; Eberl, H.C.; et al. Design of amidobenzimidazole STING receptor agonists with systemic activity. Nature 2018, 564, 439–443. [Google Scholar] [CrossRef]
- Li, X.D.; Wu, J.X.; Gao, D.X.; Wang, H.; Sun, L.J.; Chen, Z.J.J. Pivotal Roles of cGAS-cGAMP Signaling in Antiviral Defense and Immune Adjuvant Effects. Science 2013, 341, 1390–1394. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Y.; Yin, Q.; Kuss, P.; Maliga, Z.; Millan, J.L.; Wu, H.; Mitchison, T.J. Hydrolysis of 2’ 3’-cGAMP by ENPP1 and design of nonhydrolyzable analogs. Nat. Chem. Biol. 2015, 11, 235. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, P.Y.; Yu, Y.; Fu, Y.H.; Jiang, H.Y.; Lu, M.; Sun, Z.P.; Jiang, S.B.; Lu, L.; Wu, M.X. Pulmonary surfactant-biomimetic nanoparticles potentiate heterosubtypic influenza immunity. Science 2020, 367, eaau0810. [Google Scholar] [CrossRef]
- Aroh, C.; Wang, Z.H.; Dobbs, N.; Luo, M.; Chen, Z.J.; Gao, J.M.; Yan, N. Innate Immune Activation by cGMP-AMP Nanoparticles Leads to Potent and Long-Acting Antiretroviral Response against HIV-1. J. Immunol. 2017, 199, 3840–3848. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Cheng, H.; Xu, Q.M.; Tan, X.S. Encapsulation of STING Agonist cGAMP with Folic Acid-Conjugated Liposomes Significantly Enhances Antitumor Pharmacodynamic Effect. In Cancer Biotherapy and Radiopharmaceuticals; Mary Ann Liebert, Inc.: New Rochelle, NY, USA, 2021. [Google Scholar]
- Wang, J.; Li, P.Y.; Wu, M.X. Natural STING Agonist as an “Ideal” Adjuvant for Cutaneous Vaccination. J. Investig. Dermatol. 2016, 136, 2183–2191. [Google Scholar] [CrossRef] [Green Version]
- Corrales, L.; Glickman, L.H.; McWhirter, S.M.; Kanne, D.B.; Sivick, K.E.; Katibah, G.E.; Woo, S.R.; Lemmens, E.; Banda, T.; Leong, J.J.; et al. Direct Activation of STING in the Tumor Microenvironment Leads to Potent and Systemic Tumor Regression and Immunity. Cell Rep. 2015, 11, 1018–1030. [Google Scholar] [CrossRef] [Green Version]
- Saito, R.; Tominaga, T. Convection-enhanced Delivery of Therapeutics for Malignant Gliomas. Neurol. Med. Chir. 2017, 57, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasnici, S.; Werner, A.; Eichhorn, M.E.; Schmitt-Sody, M.; Pahernik, S.A.; Sauer, B.; Schulze, B.; Teifel, M.; Michaelis, U.; Naujoks, K.; et al. Effect of the surface charge of liposomes on their uptake by angiogenic tumor vessels. Int. J. Cancer 2003, 105, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yu, X.; Wang, Y.; Yuan, Y.; Xiao, H.; Cheng, D.; Shuai, X. A reduction and pH dual-sensitive polymeric vector for long-circulating and tumor-targeted siRNA delivery. Adv. Mater. 2014, 26, 8217–8224. [Google Scholar] [CrossRef]
- Li, X.; Shu, C.; Yi, G.H.; Chaton, C.T.; Shelton, C.L.; Diao, J.S.; Zuo, X.B.; Kao, C.C.; Herr, A.B.; Li, P.W. Cyclic GMP-AMP Synthase Is Activated by Double-Stranded DNA-Induced Oligomerization. Immunity 2013, 39, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Sockolosky, J.T.; Dougan, M.; Ingram, J.R.; Ho, C.C.; Kauke, M.J.; Almo, S.C.; Ploegh, H.L.; Garcia, K.C. Durable antitumor responses to CD47 blockade require adaptive immune stimulation. Proc. Natl. Acad. Sci. USA 2016, 113, E2646–E2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broos, K.; Keyaerts, M.; Lecocq, Q.; Renmans, D.; Nguyen, T.; Escors, D.; Liston, A.; Raes, G.; Breckpot, K.; Devoogdt, N. Non-invasive assessment of murine PD-L1 levels in syngeneic tumor models by nuclear imaging with nanobody tracers. Oncotarget 2017, 8, 41932–41946. [Google Scholar] [CrossRef] [Green Version]
- Petrilli, R.; Eloy, J.O.; Lee, R.J.; Lopez, R.F.V. Preparation of Immunoliposomes by Direct Coupling of Antibodies Based on a Thioether Bond. Methods Mol. Biol. 2018, 1674, 229–237. [Google Scholar]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation | Particle Size (nm) | Zeta Potential (mV) | PDI | EE (%) | LE (%) |
|---|---|---|---|---|---|
| Lip/cGAMP | 175.63 ± 0.73 | −0.42 ± 1.17 | 0.185 | 83.42 ± 1.73 | 8.33 ± 0.16 |
| TVHH-Lip/cGAMP | 204.73 ± 4.21 | −13.30 ± 1.20 | 0.182 | 83.13 ± 2.58 | 8.35 ± 0.21 |
| VHH-Lip/cGAMP | 204.53 ± 3.81 | −12.80 ± 2.50 | 0.189 | 83.47 ± 6.79 | 8.34 ± 0.19 |
| Formulation | Phospholipid Concentration (mg/mL) | Protein Concentration (μg/mL) | Protein Density (μg Protein/mg Phospholipid) |
|---|---|---|---|
| TVHH-Lip/cGAMP | 11.87 ± 0.38 | 263.70 ± 32.52 | 20.20 μg/mg |
| VHH-Lip/cGAMP | 12.71 ± 0.82 | 278.72 ± 22.42 | 20.53 μg/mg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Lu, X.; Wang, X.; Ying, T.; Tan, X. Potent Therapeutic Strategies for COVID-19 with Single-Domain Antibody Immunoliposomes Neutralizing SARS-CoV-2 and Lip/cGAMP Enhancing Protective Immunity. Int. J. Mol. Sci. 2023, 24, 4068. https://doi.org/10.3390/ijms24044068
Zhou Y, Lu X, Wang X, Ying T, Tan X. Potent Therapeutic Strategies for COVID-19 with Single-Domain Antibody Immunoliposomes Neutralizing SARS-CoV-2 and Lip/cGAMP Enhancing Protective Immunity. International Journal of Molecular Sciences. 2023; 24(4):4068. https://doi.org/10.3390/ijms24044068
Chicago/Turabian StyleZhou, Yajun, Xing Lu, Xiaoqing Wang, Tianlei Ying, and Xiangshi Tan. 2023. "Potent Therapeutic Strategies for COVID-19 with Single-Domain Antibody Immunoliposomes Neutralizing SARS-CoV-2 and Lip/cGAMP Enhancing Protective Immunity" International Journal of Molecular Sciences 24, no. 4: 4068. https://doi.org/10.3390/ijms24044068