
Vaccination with Omicron Inactivated Vaccine in Pre-vaccinated Mice Protects against SARS-CoV-2 Prototype and Omicron Variants

 , , , ,
, , , ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Results
2.1. Vaccine Design and Production
2.2. Immunogenicity of the Omicron Inactivated Vaccine
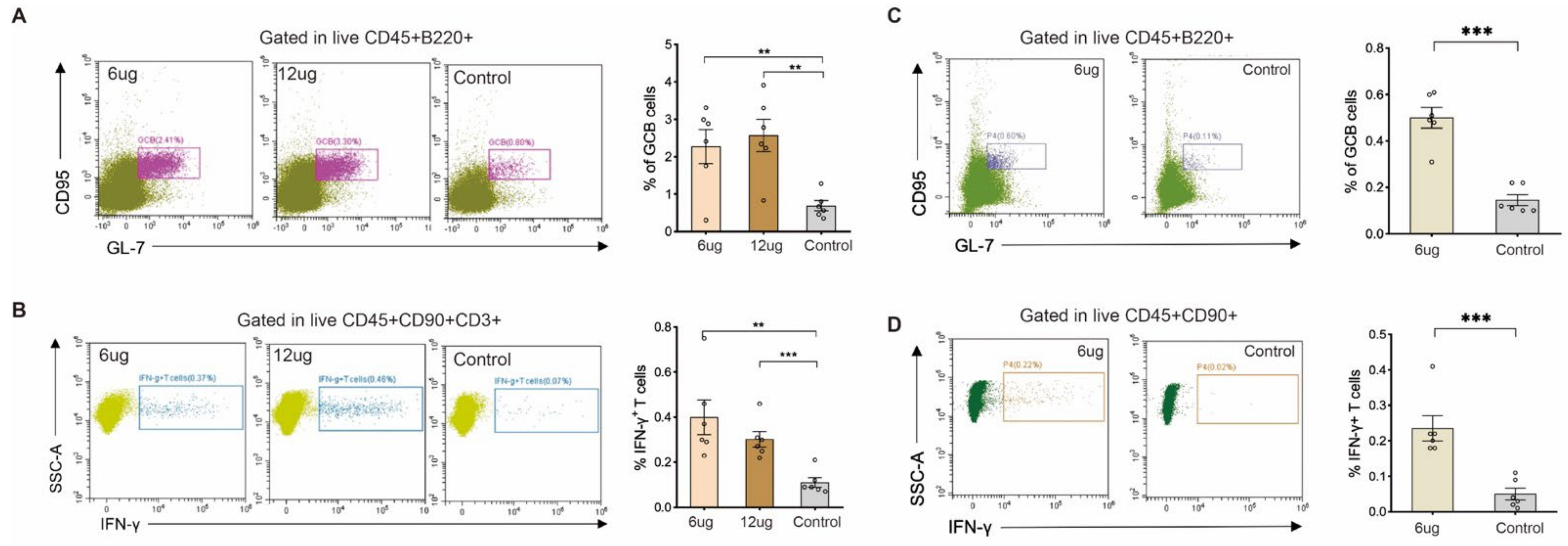
2.3. Cellular Immune Response
2.4. Heterologous Vaccination of Boosting Strategies
2.5. Safety
3. Discussion
4. Animal Models
5. Virus
6. Method Details
6.1. Phylogenic Tree Analysis
6.2. Growth Kinetics Curve
6.3. Validation of the Inactivation
6.4. Western Blotting
6.5. Vaccine Preparation
6.6. Vaccine Immunogenicity Analysis and Neutralization Assay
6.7. T Cell Response Assay
6.8. Acute Toxicity
6.9. Allergy Study
6.10. Muscle Stimulation Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeard, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connell, A.-M.; et al. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.O.; Jette, C.A.; Abernathy, M.E.; Dam, K.A.; Esswein, S.R.; Gristick, H.B.; Malyutin, A.G.; Sharaf, N.G.; Huey-Tubman, K.E.; Lee, Y.E.; et al. SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef]
- CDC. SARS-CoV-2 Variant Classifications and Definitions. 2021. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/variant-classifications.html (accessed on 23 December 2021).
- Cele, S.; Gazy, I.; Jackson, L.; Hwa, S.H.; Tegally, H.; Lustig, G.; Giandhari, J.; Pillay, S.; Wilkinson, E.; Naidoo, Y.; et al. Escape of SARS-CoV-2 501Y.V2 from neutralization by convalescent plasma. Nature 2021, 593, 142–146. [Google Scholar] [CrossRef]
- Chiu, N.-C.; Chi, H.; Tu, Y.-K.; Huang, Y.-N.; Tai, Y.-L.; Weng, S.-L.; Chang, L.; Huang, D.T.; Huang, F.-Y.; Lin, C.-Y. To mix or not to mix? A rapid systematic review of heterologous prime–boost covid-19 vaccination. Expert Rev. Vaccines 2021, 20, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- CNBC. Covid Omicron Variant Poses ‘Very High’ Global Risk and Is Likely to Spread, WHO Warns. 2021. Available online: https://www.cnbc.com/2021/11/29/who-omicron-covid-variant-poses-very-high-risk-global-spread-likely.html (accessed on 23 December 2021).
- Combadiere, B. Adaptive immunity against SARS-CoV-2. Med. Sci. 2020, 36, 908–913. [Google Scholar]
- Costa Clemens, S.A.; Weckx, L.; Clemens, R.; Almeida Mendes, A.V.; Ramos Souza, A.; Silveira, M.B.V.; Da Guarda, S.N.F.; De Nobrega, M.M.; De Moraes Pinto, M.I.; Gonzalez, I.G.S.; et al. Heterologous versus homologous COVID-19 booster vaccination in previous recipients of two doses of CoronaVac COVID-19 vaccine in Brazil (RHH-001): A phase 4, non-inferiority, single blind, randomised study. Lancet 2022, 399, 521–529. [Google Scholar] [CrossRef]
- Gao, S.J.; Guo, H.; Luo, G. Omicron variant (B.1.1.529) of SARS-CoV-2, a global urgent public health alert! J. Med. Virol. 2022, 94, 1255–1256. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Beltran, W.F.; Lam, E.C.; Astudillo, M.G.; Yang, D.; Miller, T.E.; Feldman, J.; Hauser, B.M.; Caradonna, T.M.; Clayton, K.L.; Nitido, A.D.; et al. COVID-19-neutralizing antibodies predict disease severity and survival. Cell 2021, 184, 476–488.e11. [Google Scholar] [CrossRef]
- Greaney, A.J.; Loes, A.N.; Crawford, K.H.D.; Starr, T.N.; Malone, K.D.; Chu, H.Y.; Bloom, J.D. Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe 2021, 29, 463–476.e6. [Google Scholar]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Ledford, H. Could mixing COVID vaccines boost immune response? Nature 2021, 590, 375–376. [Google Scholar] [CrossRef]
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of Epidemiological Lineages in an Emerging Pandemic Using the Pangolin Tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Stern, P.L. Key steps in vaccine development. Ann. Allergy Asthma Immunol. 2020, 125, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Quan, Y.; Xin, Z.-T.; Wrammert, J.; Ma, M.-J.; Lv, H.; Wang, T.-B.; Yang, H.; Richardus, J.H.; Liu, W.; et al. Lack of Peripheral Memory B Cell Responses in Recovered Patients with Severe Acute Respiratory Syndrome: A Six-Year Follow-Up Study. J. Immunol. 2011, 186, 7264–7268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, A.; Widera, M.; Grikscheit, K.; Toptan, T.; Schenk, B.; Pallas, C.; Metzler, M.; Kohmer, N.; Hoehl, S.; Helfritz, F.A.; et al. Reduced Neutralization of SARS-CoV-2 Omicron Variant by Vaccine Sera and Monoclonal Antibodies. medRxiv 2022. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, H.; Ye, B.; Zhao, M.; Zhan, J.; Dong, S.; Guo, Y.; Zhao, Y.; Li, M.; Liu, S.; et al. One-year sustained cellular and humoral immunities of COVID-19 convalescents. Clin. Infect. Dis. 2021, ciab884. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Dejnirattisai, W.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.E.; Tuekprakhon, A.; Nutalai, R.; et al. Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. Cell 2021, 184, 2348–2361.e6. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent | Dosage |
|---|---|
| Purified anti-mouse CD16/32 Antibody | 0.125 μg/50 μL |
| BV421 anti-mouse CD45 Antibody | 0.05 μg/50 μL |
| PE/Cyanine7 anti-mouse CD3ε Antibody | 0.05 μg/50 μL |
| FITC anti-mouse CD90.2 Antibody | 0.05 μg/50 μL |
| AF700 anti-mouse/human CD45R/B220 Antibody | 0.05 μg/50 μL |
| Super Bright 600 anti-mouse CD4 Antibody | 0.05 μg/50 μL |
| Fixable Viability Dye eFluor™ 506 | 1000× |
| FITC anti-mouse GL-7 Antibody | 0.1 μg/50 μL |
| APC anti-mouse IFN-γ Antibody | 0.15 μg/50 μL |
| PE anti-mouse CD95 Antibody | 0.1 μg/50 μL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Tan, W.; Lou, Z.; Zhao, Y.; Zhang, J.; Liang, H.; Li, N.; Zhu, X.; Ding, L.; Huang, B.; et al. Vaccination with Omicron Inactivated Vaccine in Pre-vaccinated Mice Protects against SARS-CoV-2 Prototype and Omicron Variants. Vaccines 2022, 10, 1149. https://doi.org/10.3390/vaccines10071149
Zhang Y, Tan W, Lou Z, Zhao Y, Zhang J, Liang H, Li N, Zhu X, Ding L, Huang B, et al. Vaccination with Omicron Inactivated Vaccine in Pre-vaccinated Mice Protects against SARS-CoV-2 Prototype and Omicron Variants. Vaccines. 2022; 10(7):1149. https://doi.org/10.3390/vaccines10071149
Chicago/Turabian StyleZhang, Yuntao, Wenjie Tan, Zhiyong Lou, Yuxiu Zhao, Jin Zhang, Hongyang Liang, Na Li, Xiujuan Zhu, Ling Ding, Baoying Huang, and et al. 2022. "Vaccination with Omicron Inactivated Vaccine in Pre-vaccinated Mice Protects against SARS-CoV-2 Prototype and Omicron Variants" Vaccines 10, no. 7: 1149. https://doi.org/10.3390/vaccines10071149
APA StyleZhang, Y., Tan, W., Lou, Z., Zhao, Y., Zhang, J., Liang, H., Li, N., Zhu, X., Ding, L., Huang, B., Zhou, W., Guo, Y., Yang, Z., Qiao, Y., He, Z., Ma, B., He, Y., Zhu, D., Wang, Z., ... Yang, X. (2022). Vaccination with Omicron Inactivated Vaccine in Pre-vaccinated Mice Protects against SARS-CoV-2 Prototype and Omicron Variants. Vaccines, 10(7), 1149. https://doi.org/10.3390/vaccines10071149