
Serological and Molecular Investigation of SARS-CoV-2 in Horses and Cattle in Switzerland from 2020 to 2022
 , , , , ,
, , , , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Collection
2.2. SARS-CoV-2 Immunoassays
2.2.1. Enzyme-Linked Immunosorbent Assay (ELISA)
2.2.2. Surrogate Virus Neutralization Test
2.2.3. Pseudotype-Based Virus Neutralization Assay
2.2.4. Indirect Immunofluorescence Test
2.2.5. Assessment of Cross-Reactivity to Bovine Coronavirus
2.3. Molecular Analysis
2.4. Statistics
3. Results
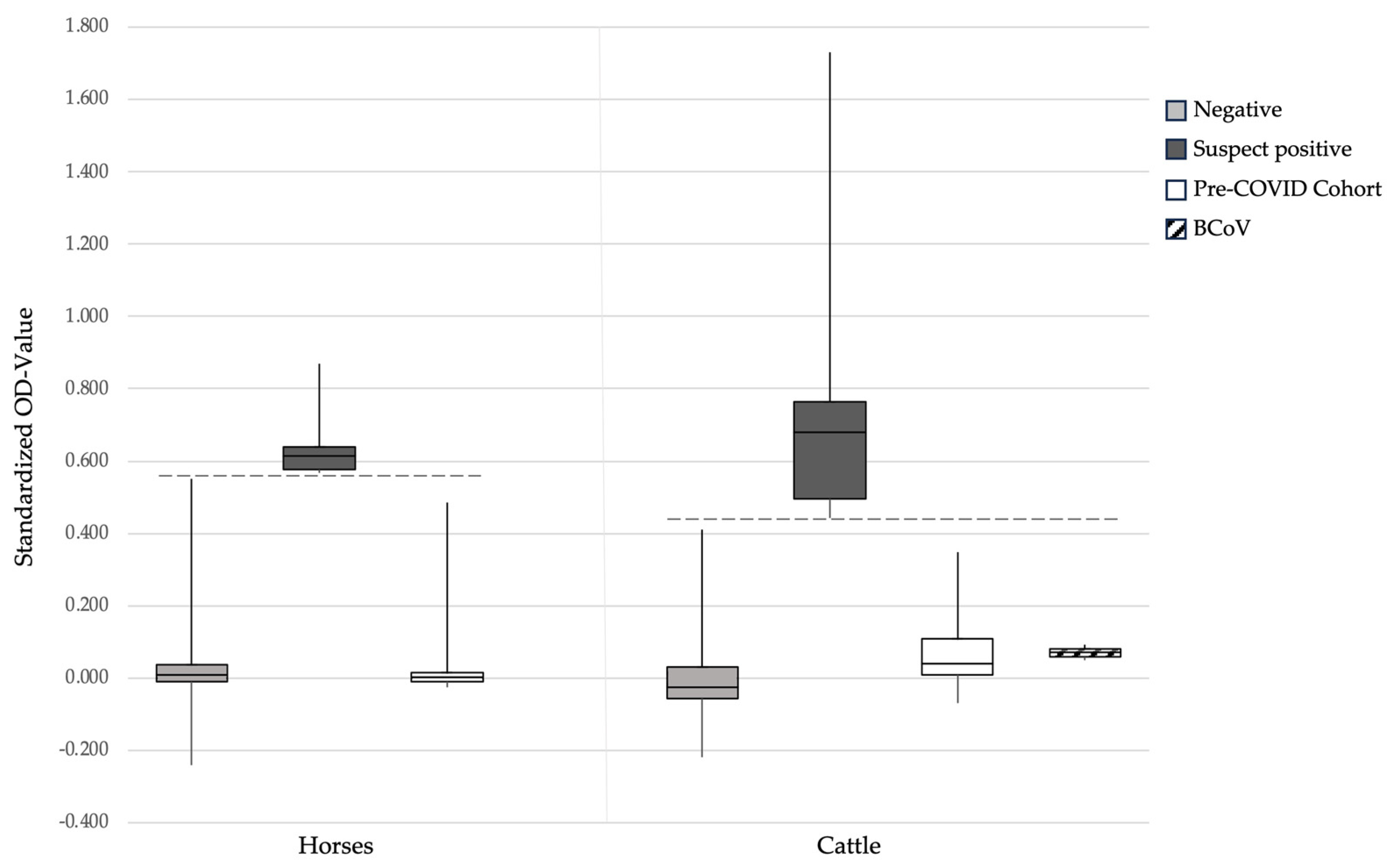
3.1. SARS-CoV-2 RBD-ELISA
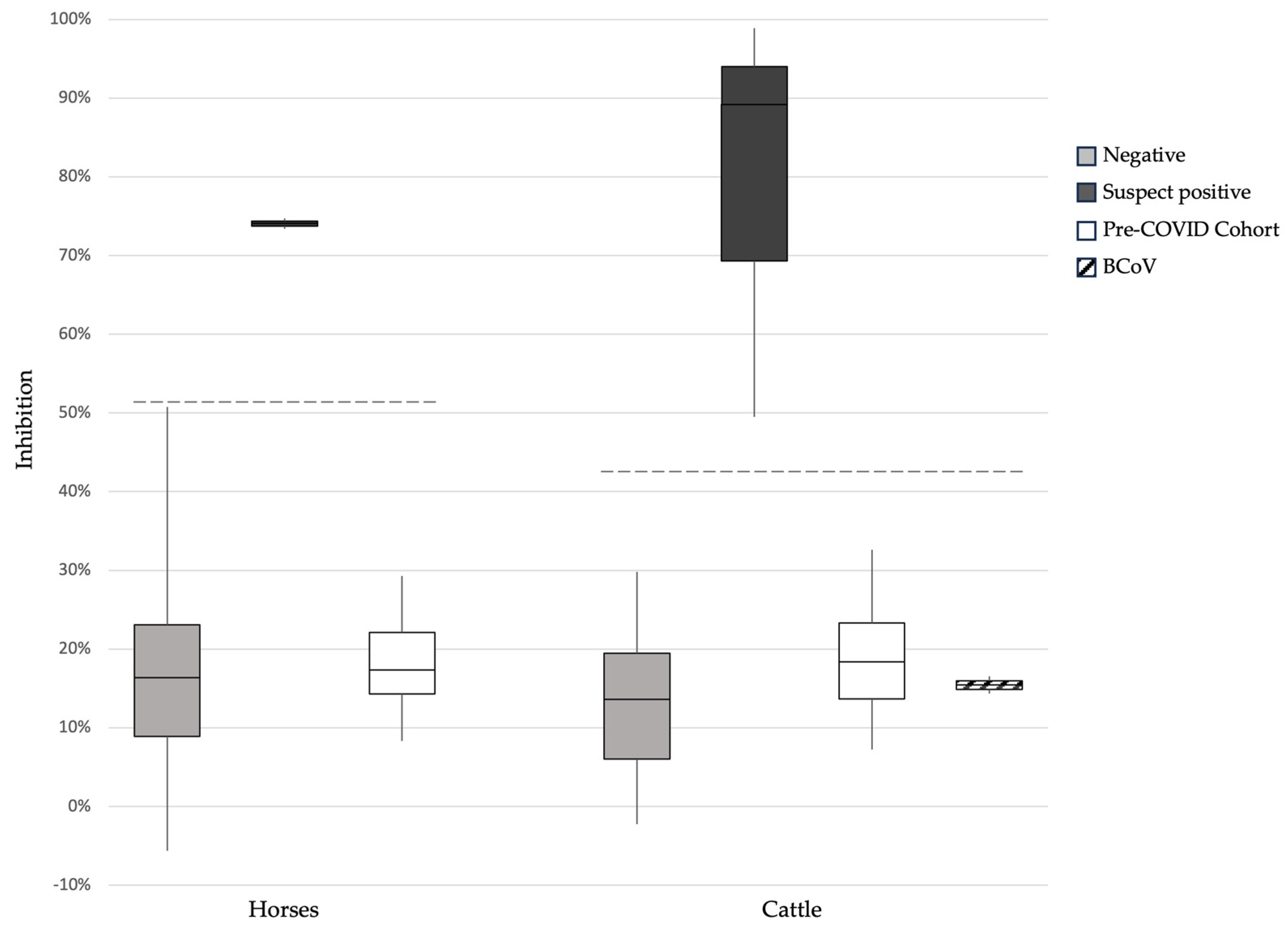
3.2. Surrogate Virus Neutralization Test
3.3. Pseudotype-Based Neutralization Assay
3.4. Immunofluorescence Test
3.5. Assessment of Potential Cross-Reactivity between SARS-CoV-2 and the Bovine Coronavirus in RBD-ELISA and sVNT
3.6. Molecular Analysis
3.7. Summary of All Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WOAH. COVID-19—Events in Animals. Available online: https://www.woah.org/en/what-we-offer/emergency-preparedness/covid-19/#ui-id-3 (accessed on 13 December 2023).
- vetmeduni. Coplexity Science Hub Vienna, SARS-ANI VIS: A Global Open Access Dataset of Reported SARS-CoV-2 Events in Animals. Available online: https://vis.csh.ac.at/sars-ani/#overview (accessed on 13 December 2023).
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef]
- Feng, A.; Bevins, S.; Chandler, J.; DeLiberto, T.J.; Ghai, R.; Lantz, K.; Lenoch, J.; Retchless, A.; Shriner, S.; Tang, C.Y.; et al. Transmission of SARS-CoV-2 in free-ranging white-tailed deer in the United States. Nat. Commun. 2023, 14, 4078. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.; Smith, D.; Ghai, R.R.; Wallace, R.M.; Torchetti, M.K.; Loiacono, C.; Murrell, L.S.; Carpenter, A.; Moroff, S.; Rooney, J.A.; et al. First Reported Cases of SARS-CoV-2 Infection in Companion Animals—New York, March–April 2020. MMWR Morb. Mortal Wkly. Rep. 2020, 69, 710–713. [Google Scholar] [CrossRef] [PubMed]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef]
- Luan, J.; Lu, Y.; Jin, X.; Zhang, L. Spike protein recognition of mammalian ACE2 predicts the host range and an optimized ACE2 for SARS-CoV-2 infection. Biochem. Biophys. Res. Commun. 2020, 526, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.P.; Pfenning, A.R.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef] [PubMed]
- Di Teodoro, G.; Valleriani, F.; Puglia, I.; Monaco, F.; Di Pancrazio, C.; Luciani, M.; Krasteva, I.; Petrini, A.; Marcacci, M.; D’Alterio, N.; et al. SARS-CoV-2 replicates in respiratory ex vivo organ cultures of domestic ruminant species. Vet. Microbiol. 2021, 252, 108933. [Google Scholar] [CrossRef] [PubMed]
- Lean, F.Z.X.; Nunez, A.; Spiro, S.; Priestnall, S.L.; Vreman, S.; Bailey, D.; James, J.; Wrigglesworth, E.; Suarez-Bonnet, A.; Conceicao, C.; et al. Differential susceptibility of SARS-CoV-2 in animals: Evidence of ACE2 host receptor distribution in companion animals, livestock and wildlife by immunohistochemical characterisation. Transbound. Emerg. Dis. 2022, 69, 2275–2286. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Lauth, A.M.; Walker, A.; Guilbert, L.; Porter, S.; Hartwig, A.; McVicker, E.; Bielefeldt-Ohmann, H.; Bowen, R.A. Susceptibility of livestock to SARS-CoV-2 infection. Emerg. Microbes Infect. 2021, 10, 2199–2201. [Google Scholar] [CrossRef]
- Lawton, K.O.Y.; Arthur, R.M.; Moeller, B.C.; Barnum, S.; Pusterla, N. Investigation of the Role of Healthy and Sick Equids in the COVID-19 Pandemic through Serological and Molecular Testing. Animals 2022, 12, 614. [Google Scholar] [CrossRef]
- Pusterla, N.; Chaillon, A.; Ignacio, C.; Smith, D.M.; Barnum, S.; Lawton, K.O.Y.; Smith, G.; Pickering, B. SARS-CoV-2 Seroconversion in an Adult Horse with Direct Contact to a COVID-19 Individual. Viruses 2022, 14, 1047. [Google Scholar] [CrossRef]
- Lawton, K.; Keller, S.M.; Barnum, S.; Arredondo-Lopez, C.; Spann, K.; Pusterla, N. Seroprevalence of SARS-CoV-2 in 1186 Equids Presented to a Veterinary Medical Teaching Hospital in California from 2020 to 2022. Viruses 2022, 14, 2497. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.C.; Bevins, S.N.; Ellis, J.W.; Linder, T.J.; Tell, R.M.; Jenkins-Moore, M.; Root, J.J.; Lenoch, J.B.; Robbe-Austerman, S.; DeLiberto, T.J.; et al. SARS-CoV-2 exposure in wild white-tailed deer (Odocoileus virginianus). Proc. Natl. Acad. Sci. USA 2021, 118, e2114828118. [Google Scholar] [CrossRef]
- Cool, K.; Gaudreault, N.N.; Morozov, I.; Trujillo, J.D.; Meekins, D.A.; McDowell, C.; Carossino, M.; Bold, D.; Mitzel, D.; Kwon, T.; et al. Infection and transmission of ancestral SARS-CoV-2 and its alpha variant in pregnant white-tailed deer. Emerg. Microbes Infect. 2022, 11, 95–112. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental Infection of Cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979–2981. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Böttcher, J.; Amelung, S.; Albrecht, K.; Gärtner, T.; Donat, K.; Beer, M. Antibodies against SARS-CoV-2 Suggestive of Single Events of Spillover to Cattle, Germany. Emerg. Infect. Dis. 2022, 28, 1916–1918. [Google Scholar] [CrossRef]
- Fiorito, F.; Iovane, V.; Pagnini, U.; Cerracchio, C.; Brandi, S.; Levante, M.; Marati, L.; Ferrara, G.; Tammaro, V.; De Carlo, E.; et al. First Description of Serological Evidence for SARS-CoV-2 in Lactating Cows. Animals 2022, 12, 1459. [Google Scholar] [CrossRef]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Klaus, J.; Zini, E.; Hartmann, K.; Egberink, H.; Kipar, A.; Bergmann, M.; Palizzotto, C.; Zhao, S.; Rossi, F.; Franco, V.; et al. SARS-CoV-2 Infection in Dogs and Cats from Southern Germany and Northern Italy during the First Wave of the COVID-19 Pandemic. Viruses 2021, 13, 1453. [Google Scholar] [CrossRef]
- Embregts, C.W.E.; Verstrepen, B.; Langermans, J.A.M.; Boszormenyi, K.P.; Sikkema, R.S.; de Vries, R.D.; Hoffmann, D.; Wernike, K.; Smit, L.A.M.; Zhao, S.; et al. Evaluation of a multi-species SARS-CoV-2 surrogate virus neutralization test. One Health 2021, 13, 100313. [Google Scholar] [CrossRef]
- Tyson, G.B.; Jones, S.; Montreuil-Spencer, C.; Logan, N.; Scott, S.; Sasvari, H.; McDonald, M.; Marshall, L.; Murcia, P.R.; Willett, B.J.; et al. Increase in SARS-CoV-2 Seroprevalence in UK Domestic Felids Despite Weak Immunogenicity of Post-Omicron Variants. Viruses 2023, 15, 1661. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmeier, E.; Chan, T.; Klaus, J.; Pineroli, B.; Geisser, E.; Hofmann-Lehmann, R.; Meli, M.L. A Pre- and Within-Pandemic Survey of SARS-CoV-2 RNA in Saliva Swabs from Stray Cats in Switzerland. Viruses 2022, 14, 681. [Google Scholar] [CrossRef] [PubMed]
- Krentz, D.; Zwicklbauer, K.; Felten, S.; Bergmann, M.; Dorsch, R.; Hofmann-Lehmann, R.; Meli, M.L.; Spiri, A.M.; von Both, U.; Alberer, M.; et al. Clinical Follow-Up and Postmortem Findings in a Cat That Was Cured of Feline Infectious Peritonitis with an Oral Antiviral Drug Containing GS-441524. Viruses 2022, 14, 2040. [Google Scholar] [CrossRef] [PubMed]
- Bundesamt für Gesundheit BAG. COVID-19 Switzerland. Available online: https://www.covid19.admin.ch/de/epidemiologic/virus-variants?variantZoomHospSeg=2022-04-10_2022-07-24 (accessed on 13 December 2023).
- Wernike, K.; Aebischer, A.; Michelitsch, A.; Hoffmann, D.; Freuling, C.; Balkema-Buschmann, A.; Graaf, A.; Müller, T.; Osterrieder, N.; Rissmann, M.; et al. Multi-species ELISA for the detection of antibodies against SARS-CoV-2 in animals. Transbound. Emerg. Dis. 2021, 68, 1779–1785. [Google Scholar] [CrossRef]
- Decaro, N.; Grassi, A.; Lorusso, E.; Patterson, E.I.; Lorusso, A.; Desario, C.; Anderson, E.R.; Vasinioti, V.; Wastika, C.E.; Hughes, G.L.; et al. Long-term persistence of neutralizing SARS-CoV-2 antibodies in pets. Transbound. Emerg. Dis. 2022, 69, 3073–3076. [Google Scholar] [CrossRef]
- Wernike, K.; Böttcher, J.; Amelung, S.; Albrecht, K.; Gärtner, T.; Donat, K.; Beer, M. Serological screening suggests single SARS-CoV-2 spillover events to cattle. bioRxiv 2022. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample ID | Sampling Date | RBD-ELISA | sVNT | B.1 | Alpha | Delta | Omicron BA.1 | Omicron BA.2 |
|---|---|---|---|---|---|---|---|---|
| #705 | May 2020 | 0.592 | 15.9 | ≤50 | ≤50 | ≤50 | ≤50 | n.t. |
| #199 | August 2020 | 0.571 | 21.8 | ≤50 | ≤50 | ≤50 | ≤50 | n.t. |
| #61 | December 2020 | 0.640 | 12.8 | ≤50 | ≤50 | ≤50 | ≤50 | n.t. |
| #451 | January 2022 | 0.634 | 11.4 | ≤50 | ≤50 | ≤50 | ≤50 | n.t. |
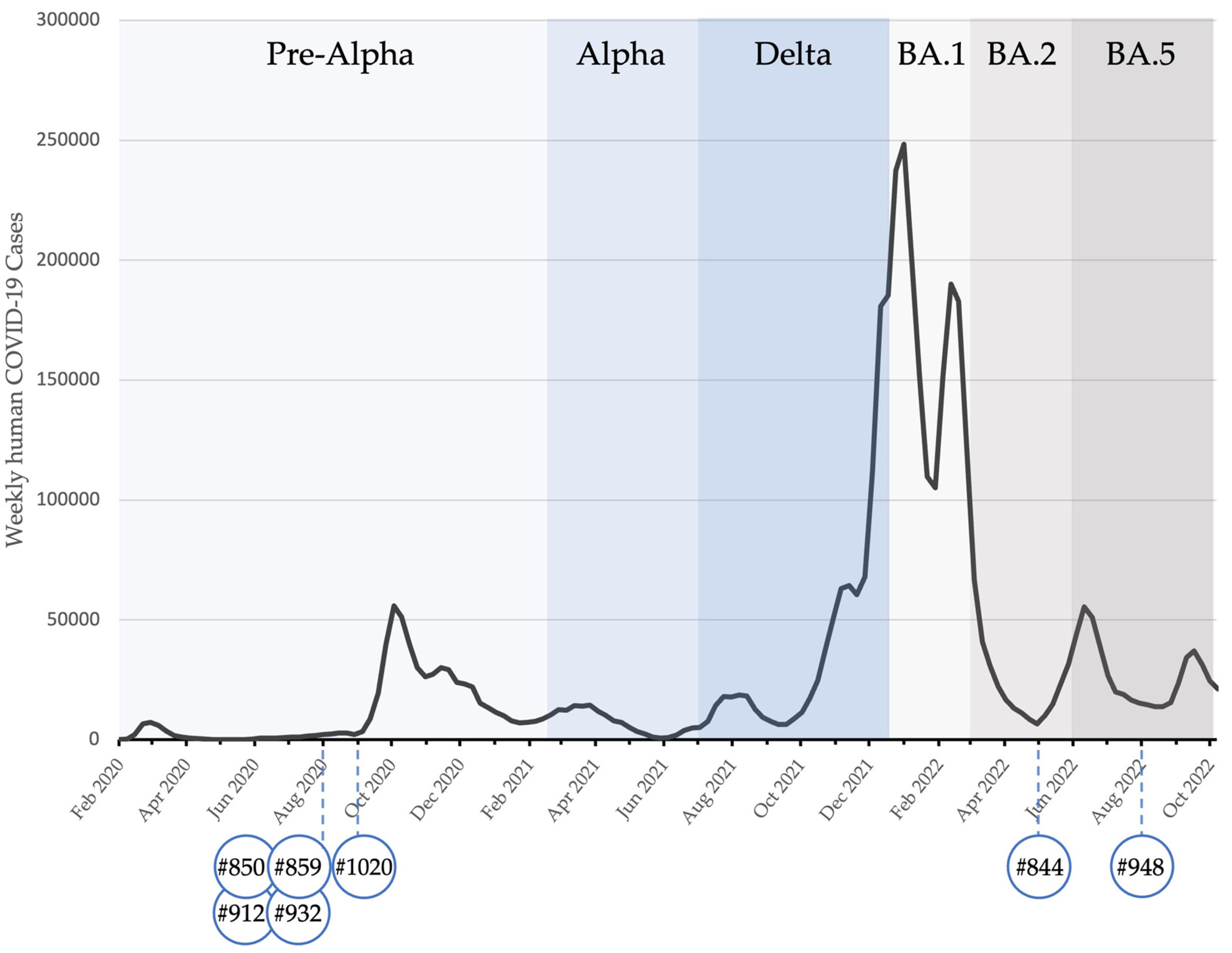
| #844 | May 2022 | 0.868 | 74.7 | 101 | 168 | 108 | 71 | 68 |
| #948 | August 2022 | 0.566 | 73.4 | 76 | 120 | 153 | 87 | 59 |
| #850 | August 2020 | 0.875 | 98.2 | 243 | 225 | 120 | ≤50 | n.t. |
| #859 | August 2020 | 1.105 | 100.6 | 543 | 371 | 207 | 71 | n.t. |
| #912 | August 2020 | 1.000 | 100.4 | 363 | 307 | 222 | 54 | n.t. |
| #932 | August 2020 | 1.097 | 100.8 | 1151 | 815 | 636 | 93 | n.t. |
| #1020 | September 2020 | 0.953 | 100.3 | 568 | 403 | 285 | 59 | n.t. |
| Sample ID | Sampling Date | RBD-ELISA | sVNT | IFA |
|---|---|---|---|---|
| #184 | March 2020 | 0.632 | −1.1 | <8 |
| #89 | March 2020 | 0.737 | −2.2 | <8 |
| #825 | April 2020 | 0.506 | 19.6 | 8 |
| #700 | April 2020 | 0.678 | 89.2 | 128 |
| #25 | June 2020 | 0.786 | 18.1 | <8 |
| #44 | June 2020 | 0.583 | 0.47 | <8 |
| #2 | June 2020 | 0.451 | 6.0 | <8 |
| #334 | July 2020 | 0.470 | 12.3 | <8 |
| #520 | October 2020 | 0.836 | 19.5 | <8 |
| #597 | June 2021 | 1.729 | 98.9 | 128 |
| #589 | June 2021 | 0.742 | 8.4 | <8 |
| #631 | November 2021 | 0.741 | 11.3 | <8 |
| #644 | December 2021 | 0.868 | 49.5 | 16 |
| #407 | April 2022 | 0.442 | 13.6 | <8 |
| #419 | April 2022 | 0.483 | 27.5 | <8 |
| Horses No. of Samples Tested | Positive | Cattle No. of Samples Tested | Positive | |
|---|---|---|---|---|
| RBD-ELISA | 1110 | 6 (suspect positive) | 830 | 15 (suspect positive) |
| sVNT | 37 | 2 | 20 | 3 |
| PVNA | 24 | 2 | n.t. | n.t. |
| IFA | n.t. | n.t. | 17 | 4 |
| BAL-PCR | 244 | 0 | n.t. | n.t. |
| Nasal RT-PCR | 67 | 0 | 16 | 0 |
| Oropharyngeal RT-PCR | 67 | 0 | 14 | 0 |
| Fecal RT-PCR | 64 | 0 | 14 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hüttl, J.; Reitt, K.; Meli, M.L.; Meili, T.; Bönzli, E.; Pineroli, B.; Ginders, J.; Schoster, A.; Jones, S.; Tyson, G.B.; et al. Serological and Molecular Investigation of SARS-CoV-2 in Horses and Cattle in Switzerland from 2020 to 2022. Viruses 2024, 16, 224. https://doi.org/10.3390/v16020224
Hüttl J, Reitt K, Meli ML, Meili T, Bönzli E, Pineroli B, Ginders J, Schoster A, Jones S, Tyson GB, et al. Serological and Molecular Investigation of SARS-CoV-2 in Horses and Cattle in Switzerland from 2020 to 2022. Viruses. 2024; 16(2):224. https://doi.org/10.3390/v16020224
Chicago/Turabian StyleHüttl, Julia, Katja Reitt, Marina L. Meli, Theres Meili, Eva Bönzli, Benita Pineroli, Julia Ginders, Angelika Schoster, Sarah Jones, Grace B. Tyson, and et al. 2024. "Serological and Molecular Investigation of SARS-CoV-2 in Horses and Cattle in Switzerland from 2020 to 2022" Viruses 16, no. 2: 224. https://doi.org/10.3390/v16020224