
Mesenchymal Stem Cell-Derived Exosomes Exhibit Promising Potential for Treating SARS-CoV-2-Infected Patients

 ,
,  ,
, 

Abstract
:1. Introduction
2. Pathological Features of COVID-19
2.1. Asymptomatic (1-2 Day of Post-Infection)
2.2. Upper Airway and Conducting Airway Response
2.3. Hypoxia and Progression to Acute Respiratory Distress Syndrome (ARDS)
3. Treatment of SARS-CoV-2 with Mesenchymal Stem Cells
4. Exosome: A Nanometer-Sized Envelope Possessing Miraculous Properties
5. Biogenesis of Exosomes
6. Mesenchymal Stem Cell-Derived Exosomes
7. Anti-Inflammatory Effect of MSC-Derived Exosomes
8. Pro-Angiogenic Role of MSC-Derived Exosomes
9. MSC-Derived Exosomes as a Prospective Treatment for COVID-19
10. Management of SARS-CoV-2-Associated Respiratory Dysfunction with Extracellular Vesicles
11. Delivery Route for Efficient Treatment against SARS-CoV-2
12. Non-EVs and Non-Vaccine-Based Treatment Regime in SARS-CoV-2
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Xing-Lou, Y.; Xian-Guang, W.; Ben, H.; Lei, Z.; Wei, Z.S.; Yan, Z.; Bei, L.; Chao-Lin, H.; Hui-Dong, C.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- De Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Coronavirus Disease (COVID-19) Dashboard. 2020. Available online: https://covid19.who.int/ (accessed on 3 March 2021).
- Trade Set to Plunge as COVID-19 Pandemic Upends Global Economy. Available online: https://www.wto.org/english/news_e/pres20_e/pr855_e.htm (accessed on 3 March 2021).
- Shan, L.; Danyi, P.; Huijun, Q.; Ke, Y.; Zhou, F.; Lin, Z. Mesenchymal stem cells as a potential therapy for COVID-19. Stem Cell Res. Ther. 2020, 11, 169. [Google Scholar]
- Mason, R.J. Pathogenesis of COVID-19 from a cell biologic perspective. Eur. Respir. J. 2020, 55, 2000607. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Du, R.-H.; Li, B.; Zheng, X.-S.; Yang, X.-L.; Hu, B.; Wang, Y.-Y.; Xiao, G.-F.; Yan, B.; Shi, Z.-L.; et al. Molecular and serological investigation of 2019-nCoV infected patients: Implication of multiple shedding routes. Emerg. Microbes Infect. 2020, 9, 386–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markus, H.; Hannah, K.W.; Nadine, K.; Marcel, M.; Christian, D.; Stefan, P. The novel coronavirus 2019 (2019-nCoV) uses the SARS-coronavirus receptor ACE2 and the cellular protease TMPRSS2 for entry into target cells. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Tortorici, M.A.; Veesler, D. Structural Insights into Coronavirus Entry; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 105, pp. 93–116. [Google Scholar]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a report of 72314 cases from the chinese center for disease control and prevention. J. Am. Med. Assoc. 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor recognition by the novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS coronavirus. J. Virol. 2020, 94, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Sims, A.C.; Baric, R.S.; Yount, B.; Burkett, S.E.; Collins, P.L.; Pickles, R.J. Severe acute respiratory syndrome coronavirus infection of human ciliated airway epithelia: Role of ciliated cells in viral spread in the conducting airways of the lungs. J. Virol. 2005, 79, 15511–15524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhom, K.; Obi, P.O.; Saleem, A. A review of exosomal isolation methods: Is Size exclusion chromatography the best option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Hancock, A.S.; Stairiker, C.J.; Boesteanu, A.C.; Monzón-Casanova, E.; Lukasiak, S.; Mueller, Y.M.; Stubbs, A.P.; García-Sastre, A.; Turner, M.; Katsikis, P.D. Transcriptome analysis of infected and bystander Type 2 alveolar epithelial cells during influenza A virus infection reveals in vivo Wntpathway downregulation. J. Virol. 2018, 92, e01325-18. [Google Scholar] [CrossRef] [Green Version]
- Mossel, E.C.; Wang, J.; Jeffers, S.; Edeen, K.E.; Wang, S.; Cosgrove, G.P.; Funk, C.J.; Manzer, R.; Miura, T.A.; Pearson, L.D.; et al. SARS-CoV replicates in primary human alveolar type II cell cultures but not in type I-like cells. Virology 2008, 372, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Viola, K.W.; Becher, A.; Tönnies, M.; Gudrun, H.; Jessica, K.; Torsten, T.B.; Paul, S.; Jens, N.; Jens, C.R.; Kolja, S.; et al. Influenza A virus’s target type II pneumocytes in the human lung. J. Infect. Dis. 2012, 206, 1685–1694. [Google Scholar]
- Qian, Z.; Travanty, E.A.; Oko, L.; Edeen, K.; Berglund, A.; Wang, J.; Ito, Y.; Holmes, K.V.; Mason, R.J. Innate immune response of human alveolar type II cells infected with severe acute respiratory syndrome–coronavirus. Am. J. Respir. Cell Mol. Biol. 2013, 48, 742–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Nikolaos, M.N.; John, G.N.; Lori, B.P.; Jason, C.G.; Vasuki, U.; Hoaxing, W.; Atsushi, S.; Kara, E.L.; Human, L.; Mitchell, R.W.; et al. Mitogenic stimulation accelerates influenza induced mortality by increasing susceptibility of alveolar type II cells to infection. Proc. Natl. Acad. Sci. USA 2017, 114, E6613–E6622. [Google Scholar] [CrossRef] [Green Version]
- de Wilde, A.H.; Snijder, E.J.; Kikkert, M. Roles of Host Gene and Non-Coding RNA Expression in Virus Infection; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 1–42. [Google Scholar]
- Sawicki, S.G.; Sawicki, D.L. Coronavirus transcription: A perspective. Curr. Top. Microbiol. Immunol. 2005, 287, 31–55. [Google Scholar] [CrossRef] [Green Version]
- Whelan, D.S.; Caplice, N.M.; Clover, A.J.P. Mesenchymal stromal cell derived CCL2 is required for accelerated wound healing. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gazdic, M.; Volarevic, V.; Arsenijevic, N.; Stojkovic, M. Mesenchymal Stem Cells: A Friend or Foe in Immune-Mediated Diseases. Stem Cell Rev. Rep. 2015, 11, 280–287. [Google Scholar] [CrossRef]
- Matthay, M.A.; Calfe, C.S.; Zhuo, H.; Thompson, B.T.; Wilson, J.G.; Levitt, J.E.; Rogers, A.J.; Gotts, J.E.; Wiener-Kronish, J.P.; Bajwa, E.K.; et al. Treatment with allogeneic mesenchymal stromal cells for moderate to severe acute respiratory distress syndrome (START study): A randomized phase 2a safety trial. Lancet Respir. Med. 2019, 7, 154–162. [Google Scholar] [PubMed]
- Xu, A.L.; Rodriguez, L.A.; Walker, K.P.; Mohammadipoor, A.; Kamucheka, R.M.; Cancio, L.C.; Batchinsky, A.I.; Antebi, B. Mesenchymal stem cells reconditioned in their own serum exhibit augmented therapeutic properties in the setting of acute respiratory distress syndrome. Stem Cells Transl. Med. 2019, 8, 1092–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, T.J.; Jackson, M.V.; Cunningham, E.K.; Kissenpfennig, A.; McAuley, D.F.; O’Kane, C.M.; Krasnodembskaya, A.D. Mesenchymal stromal cells modulate macrophages in clinically relevant lung injury models by extracellular vesicle mitochondrial transfer. Am. J. Respir. Crit. Care Med. 2017, 196, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Krasnodembskaya, A. Mesenchymal stem cell-derived extracellular vesicles for the treatment of acute respiratory distress syndrome. Stem Cells Transl. Med. 2020, 9, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 1–18. [Google Scholar] [CrossRef]
- Fujita, Y.; Kadota, T.; Araya, J.; Ochiya, T.; Kuwano, K. Clinical application of mesenchymal stem cell-derived extracellular vesicle-based therapeutics for inflammatory lung diseases. J. Clin. Med. 2018, 7, 355. [Google Scholar] [CrossRef] [Green Version]
- Lou, G.; Chen, Z.; Zheng, M.; Liu, Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp. Mol. Med. 2017, 49, e346. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.L.; Sánchez-Abarca, L.; Muntión, S.; Preciado, S.; Puig, N.; López-Ruano, G.; Hernández-Hernánde, Á.; Redondo, A.; Ortega, R.; Rodríguez, C.; et al. MSC surface markers (CD44, CD73 and CD90) can identify human MSC-derived extracellular vesicles by conventional flow cytometry. Cell Commun. Signal. 2016, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Choi, D.-Y.; Yun, S.J.; Choi, S.-M.; Kang, J.W.; Jung, J.W.; Hwang, D.; Kim, K.P.; Kim, D.-W. Proteomic analysis of microvesicles derived from human mesenchymal stem cells. J. Proteome Res. 2012, 11, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.S.; Lai, R.C.; Lee, M.M.; Choo, A.B.; Lee, C.N.; Lim, S.K. Mesenchymal stem cell secretes microparticles enriched in pre-microRNAs. Nucleic Acids Res. 2010, 38, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.C.; Tan, S.S.; Teh, B.J.; Sze, S.K.; Arslan, F.; De Kleijn, D.P.; Choo, A.; Lim, S.K. Proteolytic Potential of the MSC Exosome proteome: Implications for an exosome-mediated delivery of therapeutic proteasome. Int. J. Proteom. 2012, 2012, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chaolin, H.; Yeming, W.; Xingwang, L.; Lili, R.; Jianping, Z.; Yi, H.; Li, Z.; Guohui, F.; Jiuyang, X.; Xiaoying, G.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar]
- Williams, A.E.; Chambers, R.C. The mercurial nature of neutrophils: Still an enigma in ARDS? Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L217–L230. [Google Scholar] [CrossRef] [Green Version]
- Channappanavar, R.; Perlman, S. Pathogenic human coronavirus infections: Causes and consequences of cytokine storm and immunopathology. Semin. Immunopathol. 2017, 39, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.J.; Bermejo-Martin, J.F.; Danesh, A.; Muller, M.P.; Kelvin, D.J. Human immunopathogenesis of severe acute respiratory syndrome (SARS). Virus Res. 2008, 133, 13–19. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Feng, X.-M.; Abbott, J.; Fang, X.H.; Hao, Q.; Monsel, A.; Qu, J.-M.; Matthay, M.A.; Lee, J.W. Human mesenchymal stem cell microvesicles for treatment of Escherichia coli endotoxin-induced acute lung injury in mice. Stem Cells 2014, 32, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Abreu, S.C.; Lopes-Pacheco, M.; Weiss, D.J.; Rocco, P.R.M. Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Lung Diseases: Current Status and Perspectives. Front. Cell Dev. Biol. 2021, 9, 600711. [Google Scholar] [CrossRef]
- Park, J.; Kim, S.; Lim, H.; Liu, A.; Hu, S.; Lee, J.; Zhuo, H.; Hao, Q.; Matthay, M.A.; Lee, J.W. Therapeutic effects of human mesenchymal stem cell microvesicles in an ex vivo perfused human lung injured with severe E. coli pneumonia. Thorax 2019, 74, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Shentu, T.-P.; Huang, T.-S.; Cernelc-Kohan, M.; Chan, J.; Wong, S.S.; Espinoza, C.R.; Tan, C.; Gramaglia, I.; Van Der Heyde, H.; Chien, S.; et al. Thy-1 dependent uptake of mesenchymal stem cell-derived extracellular vesicles blocks myofibroblastic differentiation. Sci. Rep. 2017, 7, 18052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Haaften, T.; Byrne, R.; Bonnet, S.; Rochefort, G.Y.; Akabutu, J.; Bouchentouf, M.; Rey-Parra, G.J.; Galipeau, J.; Haromy, A.; Eaton, F.; et al. Airway delivery of mesenchymal stem cells prevents arrested alveolar growth in neonatal lung injury in rats. Am. J. Respir. Crit. Care Med. 2009, 180, 1131–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.; Baveja, R.; Liang, O.D.; Fernandez-Gonzalez, A.; Lee, C.; Mitsialis, S.A.; Kourembanas, S. Bone marrow stromal cells attenuate lung injury in a murine model of neonatal chronic lung disease. Am. J. Respir. Crit. Care Med. 2009, 180, 1122–1130. [Google Scholar] [CrossRef] [Green Version]
- Gomzikova, M.O.; James, V.; Rizvanov, A.A. Therapeutic application of mesenchymal stem cells derived extracellular vesicles for immunomodulation. Front. Immunol. 2019, 10, 2663. [Google Scholar] [CrossRef]
- Al-Nedawi, K.; Meehan, B.; Micallef, J.; Lhotak, V.; May, L.; Guha, A.; Rak, J.J. Intercellular transfer of the oncogenic receptor EGFR VIII by microvesicles derived from tumour cells. Nat. Cell Biol. 2008, 10, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Halkein, J.; Tabruyn, S.P.; Ricke-Hoch, M.; Haghikia, A.; Nguyen, N.-Q.-N.; Scherr, M.; Castermans, K.; Malvaux, L.; Lambert, V.; Thiry, M.; et al. MicroRNA-146a is a therapeutic target and biomarker for peripartum cardiomyopathy. J. Clin. Investig. 2013, 123, 2143–2154. [Google Scholar] [CrossRef]
- Mara, F.R.; Hongyan, Z.; Ronald, W. Exosomes function in pro- and anti-angiogenesis. Curr. Angiogenes 2013, 2, 54–59. [Google Scholar]
- Liang, X.; Zhang, L.; Wang, S.; Han, Q.; Zhao, R.C. Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 2016, 129, 2182–2189. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Chen, Y.; Chen, Y.; Meng, Q.; Sun, J.; Shao, L.; Yu, l.; Huang, H.; Hu, Y.; Yang, Z.; et al. MicroRNA-132, Delivered by Mesenchymal Stem Cell-Derived Exosomes, Promote Angiogenesis in Myocardial Infarction. Stem Cells Int. 2018, 2018, 3290372. [Google Scholar] [CrossRef] [Green Version]
- Aoki, N.; Yokoyama, R.; Asai, N.; Ohki, M.; Ohki, Y.; Kusubata, K.; Heissig, B.; Hattori, K.; Nakagawa, Y.; Matsuda, T. Adipocyte-derived microvesicles are associated with multiple angiogenic factors and induce angiogenesis in vivo and in vitro. Endocrinology 2010, 151, 2567–2576. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, S.; Dong, F.; Di Vincenzo, L.; Chilian, W.; Yin, L. Effect of exosomes from mesenchymal stem cells on angiogenesis. In Mesenchymal Stem Cell Derived Exosomes; Academic Press: Cambridge, MA, USA, 2015; pp. 177–205. [Google Scholar]
- Rosenberger, L.; Ezquer, M.; Lillo-Vera, F.; Pedraza, P.L.; Ortúzar, M.I.; González, P.L.; Figueroa-Valdés, A.I.; Cuenca, J.; Ezquer, F.; Khoury, M.; et al. Stem cell exosomes inhibit angiogenesis and tumor growth of oral squamous cell carcinoma. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Svensson, K.J.; Kucharzewska, P.; Christianson, H.C.; Sköld, S.; Löfstedt, T.; Johansson, M.C.; Mörgelin, M.; Bengzon, J.; Ruf, W.; Belting, M. Hypoxia triggers a proangiogenic pathway involving cancer cell microvesicles and PAR-2–mediated heparin-binding EGF signaling in endothelial cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13147–13152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic potential of mesenchymal cell–derived miRNA-150-5p–Expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef]
- Lai, R.C.; Aslant, F.; Lee, M.M.; Sze, N.S.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taghavi-Farahabadi, M.; Mahmoudi, M.; Soudi, S.; Hashemi, S.M. Hypothesis for the management and treatment of the COVID-19-induced acute respiratory distress syndrome and lung injury using mesenchymal stem cell-derived exosomes. Med. Hypotheses 2020, 144, 109865. [Google Scholar] [CrossRef]
- Khatri, M.; Richardson, L.A.; Meulia, T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell Res. Ther. 2018, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-L.; Chen, H.-H.; Chen, K.-H.; Chiang, J.Y.; Li, Y.-C.; Lin, H.-S.; Sung, P.-H.; Yip, H.-K. Adipose-derived mesenchymal stem cell-derived exosomes markedly protected the brain against sepsis syndrome induced injury in rat. Am. J. Transl. Res. 2019, 11, 3955–3971. [Google Scholar] [PubMed]
- Song, Y.; Dou, H.; Li, X.; Zhao, X.; Li, Y.; Liu, D.; Ji, J.; Liu, F.; Ding, L.; Ni, Y.; et al. Exosomal miR-146a contributes to the enhanced therapeutic efficacy of interleukin-1β-primed mesenchymal stem cells against sepsis. Stem Cells 2017, 35, 1208–1221. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Geng, M.; Peng, Y.; Meng, L.; Lu, S. Molecular immune pathogenesis and diagnosis of COVID-19. J. Pharm. Anal. 2020, 19, 1–7. [Google Scholar] [CrossRef]
- Devaney, J.; Horie, S.; Masterson, C.; Elliman, S.; Barry, F.; O’Brien, T.; Curley, G.F.; O’Toole, D.; Laffey, J.G. Human mesenchymal stromal cells decrease the severity of acute lung injury induced by E. coli in the rat. Thorax 2015, 70, 625–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.W.; Fang, X.; Gupta, N.; Serikov, V.; Matthay, M.A. Allogeneic human mesenchymal stem cells for treatment of E. coli endotoxin-induced acute lung injury in the ex vivo perfused human lung. Proc. Natl. Acad. Sci. USA 2009, 106, 16357–16362. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The Pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020, 583, 830–833. [Google Scholar] [CrossRef]
- Mahmoudi, M.; Taghavi, F.M.; Hashemi, S.M. Exosomes: Mediators of immune regulation. Immunoregulation 2019, 2, 3–8. [Google Scholar] [CrossRef]
- El Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Monsel, A.; Zhu, Y.G.; Gennai, S. Therapeutic effects of human mesenchymal stem cell-derived microvesicles in severe pneumonia in mice. Am. J. Respir. Crit. Care Med. 2015, 192, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Seraphin, K.; Jindrich, C.; Hans, W.D.; Uberala, K. Exosomal vaccines containing the S protein of the SARS coronavirus inducehigh levels of neutralizing antibodies. Virology 2007, 362, 26–37. [Google Scholar]
- A Pilot Clinical Study on Inhalation of Mesenchymal Stem Cells Exosomes Treating Severe Novel Coronavirus Pneumonia. Available online: https://clinicaltrials.gov/ct2/show/NCT04276987 (accessed on 3 March 2021).
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes derived from bone marrow mesenchymal stem cells as treatment for severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Available online: https://www.who.int/docs/coronaviruse/covid-19-trials (accessed on 3 March 2021).
- COVID-19 Specific T Cell Derived Exosomes (CSTC-Exo). Available online: https://clinicaltrials.gov/ct2/show/NCT04389385 (accessed on 3 March 2021).
- Golchin, A.; Seyedjafari, E.; Ardeshirylajimi, A. mesenchymal stem cell therapy for COVID-19: Present or future. Stem Cell Rev. Rep. 2020, 16, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajarshi, K.; Chatterjee, A.; Ray, S. Combating COVID-19 with mesenchymal stem cell therapy. Biotechnol. Rep. 2020, 26, e00467. [Google Scholar] [CrossRef]
- Metcalfe, S.M. Mesenchymal stem cells and management of COVID-19 pneumonia. Med. Drug Discov. 2020, 5, 100019. [Google Scholar] [CrossRef] [PubMed]
- Leng, Z.; Zhu, R.; Hou, W.; Feng, Y.; Yang, Y.; Han, Q.; Shan, G.; Meng, F.; Du, D.; Wang, S.; et al. Transplantation of ACE2- mesenchymal stem cells improves the outcome of patients with COVID-19 pneumonia. Aging Dis. 2020, 11, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Chen, J.; Li, T.; Wu, H.; Yang, W.; Li, Y.; Li, J.; Yu, C.; Nie, F.; Ma, Z.; et al. Clinical remission of a critically ill COVID-19 patient treated by human umbilical cord mesenchymal stem cells. Medicine 2020, 99, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yin, Y.; Lai, R.C.; Tan, S.S.; Choo, A.B.H.; Lim, S.K. Mesenchymal stem cells secrete immunologically active exosomes. Stem Cells Dev. 2014, 23, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noël, D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef]
- Huang, L.; Ma, W.; Ma, Y.; Feng, D.; Chen, H.; Cai, B. Exosomes in mesenchymal stem cells, a new therapeutic strategy for cardiovascular diseases? Int. J. Biol. Sci. 2015, 11, 238–245. [Google Scholar] [CrossRef]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Stella, C.; Maxime, R.; Karine, T.; Christian, J.; Danièle, N. Mesenchymal stem cell-derived exosomes improve the microenvironment of infarcted myocardium contributing to angiogenesis and anti-inflammation. Cell. Physiol. Biochem. 2015, 37, 2415–2424. [Google Scholar]
- Yu, B.; Zhang, X.; Li, X. Exosomes derived from mesenchymal stem cells. Int. J. Mol. Sci. 2014, 15, 4142–4157. [Google Scholar] [CrossRef] [Green Version]
- Maumus, M.; Jorgensen, C.; Noël, D. Mesenchymal stem cells in regenerative medicine applied to rheumatic diseases: Role of secretome and exosomes. Biochimie 2013, 95, 2229–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenis, R.; Cifù, A.; Quaglia, S.; Pistis, C.; Moretti, M.; Vicario, A.; Parodi, P.C.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; et al. Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells-derived exosomes. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, R.; Liu, T.; Yang, L.; Yin, G.; Xie, Q. Immunomodulatory effects of mesenchymal stromal cells-derived exosome. Immunol. Res. 2016, 64, 831–840. [Google Scholar]
- Chowdhury, R.; Webber, J.P.; Gurney, M.; Mason, M.D.; Tabi, Z.; Clayton, A. Cancer exosomes trigger mesenchymal stem cell differentiation into pro-angiogenic and pro-invasive myofibroblasts. Oncotarget 2014, 6, 715–731. [Google Scholar] [CrossRef]
- Gong, M.; Yu, B.; Wang, J.; Wang, Y.; Liu, M.; Paul, C.; Millard, R.W.; Xiao, D.-S.; Ashraf, M.; Xu, M. Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget 2017, 8, 45200–45212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asgarpour, K.; Shojaei, Z.; Amiri, F.; Ai, J.; Mahjoubin-Tehran, M.; Ghasemi, F.; Mirzaei, H.; ArefNezhad, R.; Hamblin, M.R. Exosomal microRNAs derived from mesenchymal stem cells: Cell-to-cell messages. Cell Commun. Signal. 2020, 18, 149. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Park, S.R.; Jung, B.K.; Jeon, Y.K.; Lee, Y.S.; Kim, M.K.; Kim, Y.G.; Jang, J.Y.; Kim, C.W. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS ONE 2013, 8, e84256. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment. Semin. Immunol. 2018, 35, 69–79. [Google Scholar] [CrossRef]
- Vrijsen, K.R.; Maring, J.A.; Chamuleau, S.A.J.; Verhage, V.; Mol, E.A.; Deddens, J.C.; Metz, C.H.G.; Lodder, K.; Van Eeuwijk, E.C.M.; Van Dommelen, S.M.; et al. Exosomes from cardiomyocyte progenitor cells and mesenchymal stem cells stimulate angiogenesis via EMMPRIN. Adv. Healthc. Mater. 2016, 5, 2555–2565. [Google Scholar] [CrossRef]
- Helia, N.; Mohit, I.; Sarah, R.; Azam, S.; Ensiyeh, H.S.; Nasser, A.; Hossein, B. Exosomes secreted by hypoxic cardiosphere-derived cells enhance tube formation and increase pro-angiogenic miRNA. J. Cell. Biochem. 2018, 119, 4150–4160. [Google Scholar]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Van Badiavas, E. Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis In Vitro. Stem Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef]
- Hu, G.-W.; Li, Q.; Niu, X.; Hu, B.; Liu, J.; Zhou, S.-M.; Guo, S.-C.; Lang, H.-L.; Zhang, C.-Q.; Wang, Y.; et al. Exosomes secreted by human-induced pluripotent stem cell-derived mesenchymal stem cells attenuate limb ischemia by promoting angiogenesis in mice. Stem Cell Res. Ther. 2015, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Ren, J.; Bai, Y.; Pei, X.; Han, Y. Exosomes from hypoxia-treated human adipose-derived mesenchymal stem cells enhance angiogenesis through VEGF/VEGF-R. Int. J. Biochem. Cell Biol. 2019, 109, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yao, K.; Huuskes, B.M.; Shen, H.H.; Zhuang, J.; Godson, C.; Brennan, E.P.; Wilkinson-Berka, J.L.; Wise, A.F.; Ricardo, S.D. Mesenchymal stem cells deliver exogenous MicroRNA-let7c via exosomes to attenuate renal fibrosis. Mol. Ther. 2016, 24, 1290–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.; Zhang, Q.; Cai, X.; Li, F.; Ma, Z.; Xu, M.; Lu, L. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J. Cell. Mol. Med. 2017, 21, 2491–2502. [Google Scholar] [CrossRef] [PubMed]
- Tseliou, E.; Fouad, J.; Reich, H.; Slipczuk, L.; De Couto, G.; Aminzadeh, M.; Middleton, R.; Valle, J.; Weixin, L.; Marbán, E. Fibroblasts rendered antifibrotic, antiapoptotic, and angiogenic by priming with cardiosphere-derived extracellular membrane vesicles. J. Am. Coll. Cardiol. 2015, 66, 599–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, A.T.J.; Toupet, K.; Bony, C.; Pirot, N.; Vozenin, M.-C.; Petit, B.; Roger, P.; Batteux, F.; Le Quellec, A.; Jorgensen, C.; et al. Antifibrotic, antioxidant, and immunomodulatory effects of mesenchymal stem cells in HOCL-induced systemic sclerosis. Arthritis Rheumatol. 2016, 68, 1013–1025. [Google Scholar] [CrossRef]
- Rong, X.; Liu, J.; Yao, X.; Jiang, T.; Wang, Y.; Xie, F. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the Wnt/β-catenin pathway. Stem Cell Res. Ther. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, X.; Liu, J.; Yao, X.; Jiang, T.; Wang, Y.; Xie, F. Immunomodulatory potential of human adipose mesenchymal stem cells derived exosomes on in vitro stimulated T cells. Front. Immunol. 2014, 5, 1–9. [Google Scholar]
- Willis, G.R.; Fernandez-Gonzalez, A.; Anastas, J.; Vitali, S.H.; Liu, X.; Ericsson, M.; Kwong, A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal stromal cell exosomes ameliorate experimental bronchopulmonary dysplasia and restore lung function through macrophage immunomodulation. Am. J. Respir. Crit. Care Med. 2018, 197, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Peng, Y.; Liu, D.; Weizmann, Y.; Mahato, R.I. Mesenchymal stem cell and derived exosome as small RNA carrier and Immunomodulator to improve islet transplantation. J. Control. Release 2016, 238, 166–175. [Google Scholar] [CrossRef]
- Casado, J.G.; Blázquez, R.; Vela, F.J.; Álvarez, V.; Tarazona, R.; Sánchez-Margallo, F.M. Mesenchymal stem cell-derived exosomes: Immunomodulatory evaluation in an antigen-induced synovitis porcine model. Front. Veter. Sci. 2017, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Nojehdehi, S.; Soudi, S.; Hesampour, A.; Rasouli, S.; Soleimani, M.; Hashemi, S.M. Immunomodulatory effects of mesenchymal stem cell–derived exosomes on experimental type-1 autoimmune diabetes. J. Cell. Biochem. 2018, 119, 9433–9443. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, J.; Liu, M.; Hu, X.; Xu, J. Study of immunomodulatory function of exosomes derived from human umbilical cord mesenchymal stem cells. Zhonghua Yi Xue Za Zhi 2015, 95, 2630–2633. [Google Scholar]
- Showalter, M.R.; Wancewicz, B.; Fiehn, O.; Archard, J.A.; Clayton, S.; Wagner, J.; Deng, P.; Halmai, J.; Fink, K.D.; Bauer, G.; et al. Primed mesenchymal stem cells package exosomes with metabolites associated with immunomodulation. Biochem. Biophys. Res. Commun. 2019, 512, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Börger, V.; Bremer, M.; Ferrer-Tur, R.; Gockeln, L.; Stambouli, O.; Becic, A.; Giebel, B. Mesenchymal stem/stromal cell-derived extracellular vesicles and their potential as novel immunomodulatory therapeutic agents. Int. J. Mol. Sci. 2017, 18, 1450. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Deregibus, M.C.; Camussi, G. The secretome of mesenchymal stromal cells: Role of extracellular vesicles in immunomodulation. Immunol. Lett. 2015, 168, 154–158. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus–infected pneumonia in Wuhan, China. J. Am. Med. Assoc. 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Tian, S.; Hu, W.; Niu, L.; Liu, H.; Xu, H.; Xiao, S.-Y. Pulmonary Pathology of Early-Phase 2019 Novel Coronavirus (COVID-19) Pneumonia in two patients with lung cancer. J. Thorac. Oncol. 2020, 15, 700–704. [Google Scholar] [CrossRef]
- Tomashefski, J.F., Jr.; Davies, P.; Boggis, C.; Greene, R.; Zapol, W.M.; Reid, L.M. The pulmonary vascular lesions of the adult respiratory distress syndrome. Am. J. Pathol. 1983, 112, 112–126. [Google Scholar] [PubMed]
- Laffey, J.G.; Matthay, M.A. Fifty years of research in ARDS. Cell-based therapy for acute respiratory distress syndrome. Biology and potential therapeutic value. Am. J. Respir. Crit. Care Med. 2017, 196, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Shah, T.G.; Predescu, D.; Predescu, S. Mesenchymal stem cells-derived extracellular vesicles in acute respiratory distress syndrome: A review of current literature and potential future treatment options. Clin. Transl. Med. 2019, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, T.; Qin, S.; Vashi, M.; Predescu, D.N.; Jeganathan, N.; Bardita, C.; Ganesh, B.; DiBartolo, S.; Fogg, L.F.; Balk, R.A.; et al. Alk5/Runx1 signaling mediated by extracellular vesicles promotes vascular repair in acute respiratory distress syndrome. Clin. Transl. Med. 2018, 7, 19. [Google Scholar] [CrossRef]
- Lai, C.P.; Mardini, O.; Ericsson, M.; Prabhakar, S.; Maguire, C.A.; Chen, J.W.; Tannous, B.A.; Breakefield, X.O. Dynamic biodistribution of extracellular vesicles in vivo using a multimodal imaging reporter. ACS Nano 2014, 8, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y.; Takeshita, F.; Kuwano, K.; Ochiya, T. RNAi Therapeutic platforms for lung diseases. Pharmaceuticals 2013, 6, 223–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinh, P.-U.C.; Paudel, D.; Brochu, H.; Popowski, K.D.; Gracieux, M.C.; Cores, J.; Huang, K.; Hensley, M.T.; Harrell, E.; Vandergriff, A.C.; et al. Inhalation of lung spheroid cell secretome and exosomes promotes lung repair in pulmonary fibrosis. Nat. Commun. 2020, 11, 1064. [Google Scholar] [CrossRef]
- Combs, J.C. SARS CoV-2: A review of current treatment regimens. Glob. Reprod. Health 2020, 5, e41. [Google Scholar] [CrossRef]


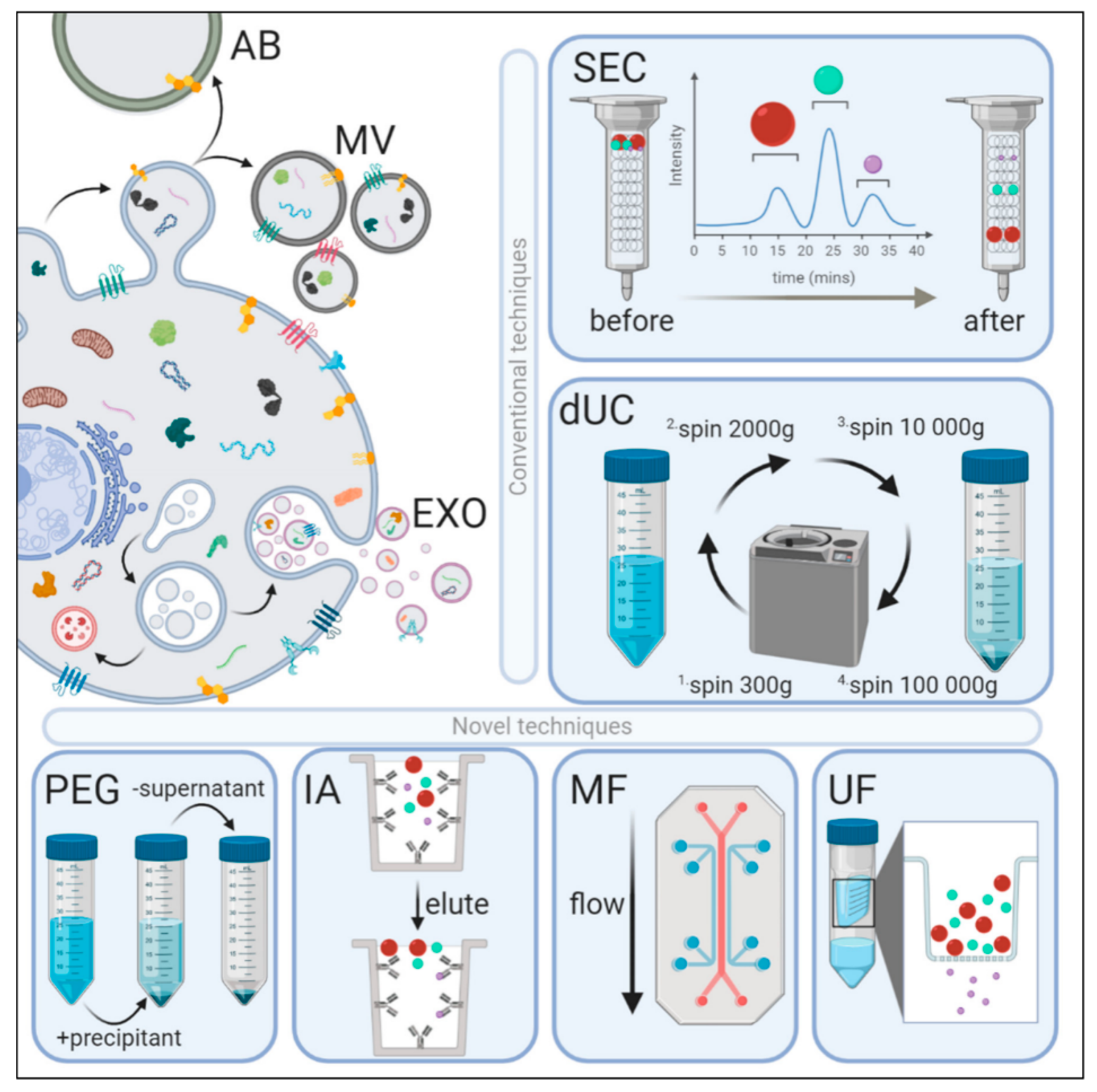{kind=link}
| Disease | Source of MSCs | Phase | First Posted Date, Initiating Company/Sponsor, Country |
|---|---|---|---|
| Chronic Autoimmune Urticaria | Auto AD-MSC | Phase 1 | 03.03.2017; Celal Bayar University, Turkey. |
| Multiple Sclerosis | Auto AD-MSC | Phase 1 Phase 2 | 01.2010; Regional University Hospital in Málaga, University Hospital Virgen Macarena, Spain. |
| 30.12.2014; American CryoStem Corporation, Cayman Islands. | |||
| AD-MSC, no specification | Phase 1 Phase 2 | 04.10.2018; Stem Cell Medicine Ltd., Israel. | |
| Auto MSC | Phase 1 Phase 2 | 21.11.2012; Karolinska Institute, Stockholm, Sweden. | |
| 16.05.2013; University of Genova, Italy. | |||
| 12.09.2014; Ottawa Hospital Research Institute, Canada. | |||
| 26.10.2010; Instituto de Salud Carlos III, Spain. | |||
| 23.12.2008; Cleveland Clinic Mellen Center, USA. | |||
| 25.05.2012; Imperial College London, United Kingdom. | |||
| UC-MSC | Phase 1 Phase 2 | 25.07.2017; Jordan University Hospital Amman, Jordan. | |
| 27.10.2015; Novo Cellular Medicine Institute LLP, India. | |||
| 16.04.2015; Genesis Limited, India. | |||
| Auto BM-MSC | Phase 1 Phase 2 | 31.03.2015; University Hospital, Toulouse, France. | |
| 18.12.2018; Karolinska Institute, Sweden. | |||
| 21.06.2011; Royan Institute, Iran. | |||
| 14.01.2014; Germans Trias i Pujol Hospital Badalona, Spain. | |||
| 13.07.2015; Vall d’Hebron Research Institute, Spain. | |||
| 10.12.2012; Andalusian Initiative for Advanced Therapies, Spain. | |||
| 29.10.2008; Hadassah Medical Organization, Israel. | |||
| 18.06.2014; Hadassah Medical Center, Israel. | |||
| 03.03.2017; Stem Cells Arabia, Jordan. | |||
| 10.07.2013; Jordan University Hospital Amman, Jordan. | |||
| 10.01.2019; Brainstorm-Cell Therapeutics, USA. | |||
| 02.11.2006; University of Cambridge, United Kingdom. | |||
| Crohn’s Disease | Allo BM-MSC | Phase 1 Phase 2 | 22.02.2012; University Hospital Liège, Belgium. |
| UC-MSC | Phase 1 Phase 2 | 04.12.2013; Kang Stem Biotech Co., Ltd., South Korea. | |
| Rheumatoid Arthritis | Auto AD-MSC | Phase 1 Phase 2 | 02.10.2018; Hope Biosciences, USA. |
| Allo AD-MSC | Phase 1 Phase 2 | 13.08.2012; TiGenix S.A.U., Spain. | |
| Auto BM-MSC | Phase 2 Phase 3 | 10.06.2013; Royan Institute, Iran. | |
| Phase 1 | 20.06.2016; Mashhad University of Medical Sciences, Iran. | ||
| 14.06.2017; MetroHealth Medical Center, USA. | |||
| 01.03.2017; Stem Cells Arabia, Jordan. | |||
| UC-MSC | Phase 1 Phase 2 | 13.01.2014; Stem Cell Institute, Panama. | |
| 15.11.2013; Translational Biosciences, Panama. | |||
| 07.03.2012; Alliancells Bioscience Corporation Limited, China. | |||
| 31.12.2015; Shenzhen Hornetcorn Bio-technology Company, LTD, China. | |||
| 04.02.2019; Baylx Inc., USA. | |||
| 07.08.2018; Kang Stem Biotech Co., Ltd., South Korea. | |||
| Systemic Lupus Erythematosus | UC-MSC in Plasma-Lyte A solution | Phase 2 | 17.12.2015; Medical University of South Carolina, USA. |
| MSCs from UC, BM, AD, and DP and other in Plasma-Lyte A solution. | Phase 1 | 31.05.2017; Medical University of South Carolina, USA. | |
| Allo BM-MSC | Phase 1 Phase 2 | 17.06.2008; Nanjing Medical University, China. | |
| UC-MSC | Phase 1 Phase 2 | 05.12.2012; Nanjing Medical University, China. | |
| 01.07.2018; Saint-Louis Hospital, France. | |||
| Lupus Nephritis | Allo BM-MSC | Phase 1 Phase 2 | 17.09.2018; University Hospital Río Hortega, Spain. |
| 02.06.2017; Corestem, Inc., South Korea. | |||
| Auto MSCs | Phase 1 Phase 2 | 16.04.2008; Organ Transplant Institute, China. | |
| UC-MSCs | Phase 2 | 09.07.2018; Lingyun Sun, China. | |
| 28.02.2012; CytoMed & Beike, China. | |||
| Type 1 Diabetes | Auto BM-MSCs | Phase 1 Phase 2 | 25.03.2011; University of Sao Paulo, Brazil. |
| 05.12.2017; Central Hospital, Nancy, France. | |||
| Auto BM-MSC | Phase 2 Phase 3 | 07.07.2010; Southwest Hospital, Third Military Medical University, China. | |
| UC-MSCs | Phase 1 Phase 2 | 13.10.2010; Stem Cell Research Center of Medical School Hospital of Qingdao University, China. | |
| 16.06.2011; Fuzhou General Hospital, China. | |||
| 2.06.2011; Shenzhen Beike Bio-Technology Co., Ltd., China. | |||
| 05.05.2016; The Affiliated Nanjing Drum Tower Hospital of Nanjing University Medical School, China. | |||
| Cotransplantation of allograft Islet and auto MSCs | Phase 1 Phase 2 | 28.03.2008; Fuzhou General Hospital, China. | |
| Auto MSCs | Phase 2 | 07.02.2014; Uppsala University Hospital, Sweden. | |
| Auto BM-MSC and Allo UC-MSC combined with PRP | Phase 1 Phase 2 | 01.04.2017; Van Hanh General Hospital University of Science, Vietnam. | |
| Allo BM-MSC | Phase 2 | 08.09.2016; Clínica Alemana de Santiago, Chile. | |
| Allo AD-MSC with auto BM-MSC | Phase 1 | 20.10.2016; Sophia Al-Adwan, University of Jordan, Jordan. | |
| Auto BM-MSC and UC-MSCs | Phase 1 | 14.06.2010; Cellonis Biotechnology Co. Ltd., China. | |
| PROCHYMAL® - adult MSCs | Phase 2 | 04.06.2008; Mesoblast International, USA. | |
| Men-MSCs | Phase 1 Phase 2 | 21.12.2011; Evans Biosciences Co., Ltd., Zhejiang University, China. | |
| Neuromyelitis Optica Spectrum Disorders | Auto MSCs | Phase 2 | 25.09.2014; Tianjin Medical University General Hospital, China. |
| Sjögren’s Syndrome | Allo MSCs | Phase 1 Phase 2 | 06.08.2009; The Affiliated Nanjing Drum Tower Hospital of Nanjing University Medical School, China. |
| Hepatitis | UC-MSCs | Phase 1 Phase 2 | 10.08.2012; Beijing 302 Hospital, China. |
| Articles | Conclusion of the Study | References |
|---|---|---|
| Mesenchymal stem cells as a potential therapy for COVID-19 | A summary of clinical trials of MSCs treatments on ALI/ARDS and raise MSCs as a hopefully alternative therapy for COVID-19. | [1] |
| Mesenchymal Stem Cell Therapy for COVID-19: Present or Future | Study of MSCs-based improvement in patient’s immunological responses to COVID-19. | [2] |
| Combating COVID-19 with mesenchymal stem cell therapy | Highlights implications associated with MSC therapy application in case of COVID-19 | [3] |
| Mesenchymal stem cells and management of COVID-19 pneumonia | Collection of studies suggesting improving patient’s biological resistance to COVID-19 using stem cells | [4] |
| Transplantation of ACE2 - Mesenchymal Stem Cells Improves the Outcome of Patients With COVID-19 Pneumonia | The intravenous transplantation of MSCs was reliable, safe and efficient for treatment of COVID-19 pneumonia, especially for the patients in critically severe condition. | [5] |
| Clinical remission of a critically ill COVID-19 patient treated by human umbilical cord mesenchymal stem cells (hUC-MSCs) | The adoptive transfer therapy of hUC-MSCs might be an ideal choice to be used or combined with other immune modulating agents to treat the critically ill COVID19 patients. | [6] |
| Articles | Conclusion of the Study | Reference |
|---|---|---|
| Mesenchymal stem cells secrete immunologically active exosomes | Infusion of MSC exosomes enhanced the survival of allogenic skin graft in mice and increased Tregs. | [1] |
| Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis | Demonstration of the therapeutic potential of MSCs-derived EVs in inflammatory arthritis. | [7] |
| Exosomes in mesenchymal stem cells, a new therapeutic strategy for cardiovascular diseases? | Recent advances about the role of exosomes in MSCs therapy for CVDs. | [8] |
| Mesenchymal stem cells-derived exosomes (Exos) and microparticles (MPs) protect cartilage and bone from degradation in osteoarthritis | MPs and Exos used related chondroprotective and anti-inflammatory function in vitro and protected mice from developing OA in vivo, suggesting that either Exos or MPs reproduced the main therapeutic effect of BM-MSCs. | [9] |
| Mesenchymal stem cell-derived exosomes improve the microenvironment of infarcted myocardium contributing to angiogenesis and anti-inflammation | Exosomes stimulate neovascularization and restrain the inflammation response, thus improving heart function after ischemic injury. | [10] |
| Exosomes derived from mesenchymal stem cells | This paper provides a general overview of MSC-exosomes as a new cell-free therapeutic paradigm. | [11] |
| Mesenchymal stem cells in regenerative medicine applied to rheumatic diseases: role of secretome and exosomes | Strategies for the development of the MSC secretome with respect to the release of extracellular vesicles that would have certain advantages over injection of living MSCs or administration of a single therapeutic factor or a combination of factors. | [12] |
| Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells-derived exosomes | Study suggests that the immunomodulatory properties of AMSCs-derived exosomes may be not constitutive, instead induced by the inflammatory microenvironment. | [13] |
| Immunomodulatory effects of mesenchymal stromal cells-derived exosome | IDO showed no significant changes in PBMCs exposed to MSCs-derived exosome. They conclude that exosomes and MSCs might differ in their immune-modulating activities and mechanisms. | [14] |
| Articles | Conclusion of the study | Reference |
|---|---|---|
| Cancer exosomes trigger mesenchymal stem cell differentiation into pro-angiogenic and pro-invasive myofibroblasts | Prostate cancer exosomes dominantly dictate a programme of MSC differentiation generating myofibroblasts with functional properties consistent with disease promotion. | [16] |
| Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis | Exosomal transfer of pro-angiogenic miRNAs plays an important role in MSC-mediated angiogenesis and stem cell-to-endothelial cell communication. | [17] |
| Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a | adMSC-Exo can transfer miR-125a to endothelial cells and promote angiogenesis by repressing DLL4. | [18] |
| Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells | MSC-derived exosomes may serve as a significant mediator of cell-to-cell communication within the tumor microenvironment and suppress angiogenesis by transferring anti-angiogenic molecules. | [19] |
| Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment | MSCs have a potential to exert anti-tumor activities, they largely provide service to the tumor using the multidirectional communication system established by exosomes in the TME. | [20] |
| Exosomes from cardiomyocyte progenitor cells and mesenchymal stem cells stimulate angiogenesis via EMMPRIN | CMPC and MSC exosomes have powerful pro-angiogenic effects, and this effect is largely mediated via the presence of EMMPRIN on exosomes. | [21] |
| Exosomes secreted by hypoxic cardiosphere-derived cells enhance tube formation and increase pro-angiogenic miRNA | Benefits of hypoxic CDC exosomes for the treatment of cardiac diseases by induction of angiogenesis via enrichment of pro-angiogenic exosomal miRNAs. | [22] |
| Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis in vitro | MSC exosomes were found to activate several signaling pathways important in wound healing (AKT, ERK, and STAT3) and induce the expression of a number of growth factors [hepatocyte growth factor (HGF), insulin-like growth factor-1 (IGF1), nerve growth factor (NGF), and stromal-derived growth factor-1 (SDF1)]. | [23] |
| Exosomes secreted by human-induced pluripotent stem cell-derived mesenchymal stem cells attenuate limb ischemia by promoting angiogenesis in mice | Implanted iMSCs-Exo was able to protect limbs from ischemic injury via the promotion of angiogenesis, which indicated that iMSCs-Exo may be a novel therapeutic approach in the treatment of ischemic diseases. | [24] |
| Exosomes from hypoxia-treated human adipose-derived mesenchymal stem cells enhance angiogenesis through VEGF/VEGF-R | Exosomes from hypoxia-treated human ADSCs possess a higher capacity to enhance angiogenesis in fat grafting, at least partially, via regulating VEGF/VEGF-R signaling. | [25] |
| Articles | Conclusion of the Study | Reference |
|---|---|---|
| Mesenchymal stem cells deliver exogenous microRNA-let7c via exosomes to attenuate renal fibrosis | The effective antifibrotic function of engineered MSCs is able to selectively transfer miR-let7c to damaged kidney cells and will pave the way for the use of MSCs for therapeutic delivery of miRNA targeted at kidney disease. | [26] |
| Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation | The effective anti-fibrotic function of engineered ADSCs is able to selectively transfer miR-181-5p to damaged liver cells and will pave the way for the use of exosome-ADSCs for therapeutic delivery of miRNA targeting liver disease. | [27] |
| Mesenchymal stem cells in regenerative medicine applied to rheumatic diseases: role of secretome and exosomes | Strategies for the development of the MSC secretome with respect to the release of extracellular vesicles that would have certain advantages over injection of living MSCs or administration of a single therapeutic factor or a combination of factors. | [12] |
| Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis | The hUC-MSC-Exo could ameliorate CCl4-induced liver fibrosis by constraining EMT and protecting hepatocytes | [103] |
| Mesenchymal stem cells-derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis | MPs and Exos used related chondroprotective and anti-inflammatory function in vitro and protected mice from using OA in vivo, suggesting that either Exos or MPs reproduced the main therapeutic effect of BM-MSCs. | [9] |
| Fibroblasts rendered antifibrotic, antiapoptotic, and angiogenic by priming with cardiosphere-derived extracellular membrane vesicles (CSp-EMVs) | CSp-EMVs alter fibroblast phenotype and secretome in a salutary positive-feedback loop. The phenotypic conversion of inert cells to therapeutically active cells uncovers a novel mechanism for amplification of exosome bioactivity. | [28] |
| Antifibrotic, antioxidant, and immunomodulatory effects of mesenchymal stem cells in HOCl-induced systemic sclerosis (SSc) | This work shows the beneficial and systemic effects of MSC administration in the HOCl murine model of diffuse SSc, which is a promising finding from a clinical perspective. | [29] |
| Human bone marrow mesenchymal stem cells-derived exosomes (hBM-MSC-Exo) alleviate liver fibrosis through the Wnt/β-catenin pathway | The hBM-MSCs-Exo treatment could ameliorate CCl4-induced liver fibrosis via inhibition of HSC activation through the Wnt/β-catenin pathway. | [30] |
| Articles | Conclusion of the Study | Reference |
|---|---|---|
| Immunomodulatory potential of human adipose mesenchymal stem cells-derived exosomes on in vitro stimulated T cells | MSCs-derived exosomes are a cell-derived product that could be considered as a therapeutic agent for the treatment of inflammation-related diseases. | [31] |
| Immunomodulatory effects of mesenchymal stromal cells-derived exosome | IDO showed no significant changes in PBMCs exposed to MSCs-derived exosome. They conclude that exosome and MSCs might differ in their immune-modulating activities and mechanisms. | [14] |
| Mesenchymal stromal cell exosomes ameliorate experimental bronchopulmonary dysplasia and restore lung function through macrophage immunomodulation | The MSC-derived exosome mechanism of action is associated with modulation of lung macrophage phenotype. | [32] |
| Mesenchymal stem cell and derived exosome as small RNA carrier and Immunomodulator to improve islet transplantation | This work elucidated the mechanisms of RNA delivery from hBMSCs to human islets and the immunosuppressive effect of hBMSC and peripheral blood mononuclear cell co-cultured exosomes for improving islet transplantation. | [33] |
| Mesenchymal stem cell-derived exosomes: immunomodulatory evaluation in an antigen-induced synovitis porcine model | The study suggested immunomodulatory effect of the exosomes and pointed out that they may represent a promising therapeutic option for the treatment of synovitis. | [34] |
| Immunomodulatory effects of mesenchymal stem cell–derived exosomes on experimental type-1 autoimmune diabetes | AD-MSC’s exosomes exert ameliorative effects on autoimmune T1DM through increasing regulatory T-cell population and their products without a change in the proliferation index of lymphocytes, which makes them more effective and practical candidates. | [35] |
| Study of immunomodulatory function of exosomes derived from human umbilical cord mesenchymal stem cells (hUC-MSCs) | The hUC-MSCs-exosome has the immunomodulatory function in vitro, which could be a new therapeutic agent for the treatment of immune disorders. | [36] |
| Primed mesenchymal stem cells package exosomes with metabolites associated with immunomodulation | MSCs exposed to priming culture conditions undergo glycolytic reprogramming, which homogenizes MSCs’ metabolomic profile | [37] |
| The secretome of mesenchymal stromal cells: role of extracellular vesicles in immunomodulation | Mesenchymal stromal cells influence the cells of the immune system. This influence is mainly due to the release of paracrine factors | [38] |
| Mesenchymal stem cells secrete immunologically active exosomes | Infusion of MSC exosomes enhanced the survival of allogenic skin graft in mice and increased Tregs. | [1] |
| Mesenchymal stem cells-derived exosomes (Exos) are more immunosuppressive than microparticles (MPs) in inflammatory arthritis | MSCs-derived MPs and Exos exerted an anti-inflammatory role on T and B lymphocytes independently of MSCs priming. | [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raghav, A.; Khan, Z.A.; Upadhayay, V.K.; Tripathi, P.; Gautam, K.A.; Mishra, B.K.; Ahmad, J.; Jeong, G.-B. Mesenchymal Stem Cell-Derived Exosomes Exhibit Promising Potential for Treating SARS-CoV-2-Infected Patients. Cells 2021, 10, 587. https://doi.org/10.3390/cells10030587
Raghav A, Khan ZA, Upadhayay VK, Tripathi P, Gautam KA, Mishra BK, Ahmad J, Jeong G-B. Mesenchymal Stem Cell-Derived Exosomes Exhibit Promising Potential for Treating SARS-CoV-2-Infected Patients. Cells. 2021; 10(3):587. https://doi.org/10.3390/cells10030587
Chicago/Turabian StyleRaghav, Alok, Zeeshan Ahmad Khan, Viabhav Kumar Upadhayay, Prashant Tripathi, Kirti Amresh Gautam, Brijesh Kumar Mishra, Jamal Ahmad, and Goo-Bo Jeong. 2021. "Mesenchymal Stem Cell-Derived Exosomes Exhibit Promising Potential for Treating SARS-CoV-2-Infected Patients" Cells 10, no. 3: 587. https://doi.org/10.3390/cells10030587